الاختلافات هي نصفي الكرة المخية من الدماغ الأماميوهي أكبر حجمًا ولها بنية أكثر تعقيدًا. تتكون الطبقة الخارجية للدماغ من طبقة منفصلة من الخلايا العصبية التي تتشكل القشرة الدماغية. في معظم الثدييات، تتكون القشرة الدماغية من عدد كبير من الخلايا العصبية التي لا تقع في طبقة متواصلة، ولكنها تشكل تلافيفات. التلافيف- هذه هي العلامة الرئيسية للدماغ المتطور. القشرة الدماغية مسؤولة عن العديد من الوظائف، بما في ذلك تشكيل ردود الفعل المشروطةوالتي لا تتشكل عند إزالة القشرة.
مثل الدماغ الأمامي، المخيخ في الثديياتمتطورة جدًا ولها أيضًا تلافيفات. يرجع هذا المستوى من تطور المخيخ إلى حقيقة أن الثدييات يجب أن تكون قادرة على تنسيق الحركات المعقدة والمتنوعة. الثدييات أيضا متطورة بشكل جيد أعضاء الحس.
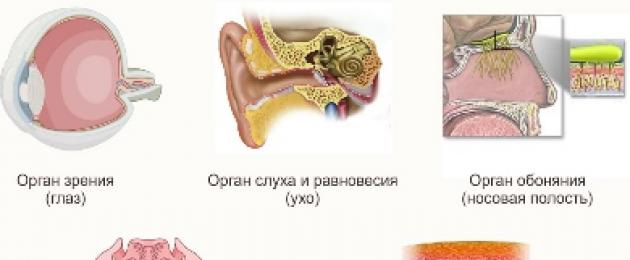
لقد تطورت الثدييات بشكل جيد أعضاء الحس: البصر والشم والسمع والذوق والشم. لكن مستوى تطور أو انخراط كل منهم في نوع معين يعتمد على موطن هذا النوع من الثدييات.

على سبيل المثال، الخلد الذي يعيش تحت الأرض لديه رؤية ضعيفة التطور. الحيتان والدلافين التي تعيش في الماء ليس لديها أي قدرة على الشم تقريبًا، على الرغم من أن العديد من الثدييات البرية لديها حاسة شم قوية جدًا. حاسة الشم الحادة ل الثدييات الأرضية- هذا هو العضو الحسي الرئيسي الذي يسمح لك بالعثور على الفريسة، والشعور باقتراب حيوان مفترس، والعثور على أنثى أو ذكر للتزاوج. يتعرف الآباء على أطفالهم من خلال الرائحة.
يلعب السمع أيضًا دورًا مهمًا بالنسبة لمعظم الثدييات. من أجل اكتشاف الأصوات الأقل وضوحًا، تمتلك الثدييات آذانالمحمول في معظمها. أيضًا، في العديد من الحيوانات التي يكون السمع مهمًا جدًا بالنسبة لها، غالبًا ما يكون سطح الأذن مغطى بشعر حساس لأدنى اهتزازات صوتية - اهتزازات.
إن رؤية الثدييات ليست حادة مثل الطيور، والعديد من الثدييات غير قادرة جزئيًا أو كليًا على تمييز الألوان. القرود فقط هي التي تستطيع التمييز بين نفس نطاق الألوان الذي يميز البشر.
أعضاء اللمس هي اهتزازاتوالتي تنمو على أجزاء حساسة من الجسم مثل الرأس. القرود، مثل البشر، تستخدم أطراف أصابعها أيضًا للمس. براعم التذوقتم تطويره بشكل جيد في الحيوانات العاشبة، بفضله يسهل تمييز النباتات الصالحة للأكل عن النباتات غير الصالحة للأكل. بسبب تعقيد الجهاز العصبي سلوك الثديياتلا يقل تعقيدا عن الطيور والحيوانات الأخرى.
سلوك الثدييات.
سلوك الثدييات معقد بسبب عدة عوامل:
الدماغ الأمامي والقشرة الدماغية المتطورة أيضًا الجهاز العصبيعمومًا؛
القدرة على إنتاج العديد ردود الفعل المشروطةطوال الحياة.
منذ الأيام الأولى من الحياة، يستطيع الأطفال التعرف على أمهم. ومع نموهم، تزداد تجربة العلاقات بين الأفراد والبيئة الخارجية، ويكتسبون المعرفة في هذه العملية ألعاب: القفز، القتال، الصيد، المطاردة، الجري، الخ). تساعد هذه الألعاب أيضًا على التطور مهارات، تحسين القدرة على التحمل لدى الأطفال. وفي المستقبل، ستساعد هذه المهارات أيضًا الثدييات عند الصيد أو إنقاذ حياتها.
تطور الثدييات طوال حياتها عددًا كبيرًا من ردود الفعل المشروطة بسبب التقلبات البيئية. ردود الفعل المشروطة القديمة التي لم يتم تعزيزها المحفزات المشروطة، يمكن أن تضيع مع مرور الوقت، مما يسمح بالاستخدام الرشيد موارد الدماغ.
§ 49. ظهور دماغ الثدييات
ظهرت أسلاف الثدييات الزواحف الصغيرة من الحطام الخشبي للعصر الكربوني مع حاسة شم متطورة وجهاز دهليزي وضعف في الرؤية ومراكز الارتباط في الدماغ المتوسط. بدأت هذه المخلوقات مسارًا تطوريًا غامضًا لم يتسم بآثار حفريات واضحة منذ ما يقرب من 60 مليون عام. فقط في أواخر العصر الترياسي ظهرت ثلاثيات الأسنان (ميجازوسترودون)،والتي يمكن اعتبارها ثدييات قديمة ولكنها متطورة بالكامل. على مدار عشرات الملايين من السنين، حدثت أحداث أدت إلى تكوين نظام ارتباطي مثالي للدماغ الأمامي، والدم الدافئ، وتطور المشيمة، وتغذية الصغار بالحليب (Kemp, 1982; Tyndale-Biscoe, Rentree) ، 1987).
دعونا نحاول تقييم التغيرات في الجهاز العصبي التي سبقت ظهور ثلاثيات الأسنان. كان لدى أسلاف الثدييات الكربونية مجموعة من الصفات المميزة لمعظم الزواحف في تلك الفترة. ولكي تصبح ثدييات، كان عليها أن تجد نفسها في بيئة توفر فيها خصائصها المورفولوجية أقصى المزايا البيولوجية.
تتمتع معظم الثدييات الحديثة بحاسة شم متطورة. يتم فقدانه بشكل ثانوي في الحيتان المسننة ويستخدمه القليل نسبيًا من قبل الململيات والخفافيش والرئيسيات. وفي حالات أخرى، تستخدم الثدييات على نطاق واسع كلاً من جهاز الشم الرئيسي والجهاز الميكعي الأنفي. بالنسبة للثدييات الأكثر بدائية، تلعب حاسة الشم دورًا رائدًا، ويمكن أن يتجاوز تمثيل مراكز المستقبلات الكيميائية في الدماغ الأمامي جميع الهياكل الأخرى مجتمعة (انظر الشكل III-19، أ). ومن الواضح أنه في المراحل الأولى من تطور الثدييات، لعبت حاسة الشم دورًا رئيسيًا. كان هذا هو السبب وراء التطور التفضيلي لنصفي الكرة الدماغية الأمامية. أدى تطور النظام الشمي إلى ظهور نصفي الكرة الدماغية الأماميين، اللذين يسيطران على بقية الدماغ. يكون حجم نصفي الكرة الأرضية المزدوج للثدييات دائمًا أكبر من حجم الهياكل الأخرى للجهاز العصبي، بغض النظر عن التخصص الذي يتمتع به نوع معين (انظر الشكل III-18؛ III-19؛ III-21؛ III-25) .
كان تطور حاسة الشم والدماغ الأمامي أول حدث عصبي كبير في تاريخ هذه المجموعة. يمكن الافتراض أن أسلاف الثدييات استخدموا الرائحة كنظام تمييزي رئيسي. تحت أي ظروف يمكن أن يحدث هذا؟ الوضع الواضح هو النشاط الليلي للثدييات القديمة، ولكن يمكن استخدام السمع والرؤية واللمس والمستقبلات الحرارية بنجاح في الصيد الليلي. فضلت الثدييات استخدام حاسة الشم، على الرغم من عدم انخفاض الحواس الأخرى بشكل كبير.
في فجر تطور الثدييات، كان هيكل الدماغ الأمامي مشابهًا لبنية دماغ ممثلي القوارض والأرنوبيات الحديثة (انظر الشكل III-18، ب؛ III-19،). أ، ب؛ III-24، أ). بحثت الثدييات القديمة عن الطعام، والشركاء الجنسيين، والفضاء المتنقل باستخدام حاسة الشم. معظم المؤلفين الذين تعاملوا مع مشاكل تطورهم يتفقون مع وجهة النظر هذه (UIinski، 1986). في هذه المرحلة من التطور، حُرم أسلاف الثدييات من الزواحف من القدرة على استخدام الحواس الأخرى بنفس فعالية الشم. على ما يبدو، كانوا يعيشون في ظلام الطبقات السفلى من ركام الكربون، حيث كانت حاسة الشم هي المستقبل البعيد الأكثر فعالية. وبالإضافة إلى حاسة الشم، يمكن أيضًا استخدام حاسة السمع واللمس هناك. كان النظام البصري ورؤية الألوان عديمي الفائدة عمليًا وفقدا خصائصهما الأصلية تدريجيًا.
ظلت الثدييات القديمة على هذه الحالة لفترة طويلة. كان هناك ما يكفي من الوقت لإنشاء اتصالات تكاملية بين المراكز القشرية التناسلية للجهاز الميكعي الأنفي والأنظمة الحسية الحركية لأجزاء أخرى من الدماغ. نشأ مركز جديد لصنع القرار على أساس البدائية القشرية المتواضعة للنظام الشمي الجنسي لدى الزواحف. ومن الواضح أنه كان يشمل في الأصل المراكز المكعية الأنفية والحركية والتذوقية.
في المرحلة الأولى من تطور الثدييات، تم تحسين الجهاز السمعي بسبب الدرنات الخلفية للرباعي التوائم. وهي أكثر تطوراً في الثدييات منها في الزواحف والطيور (انظر الشكل III-22، د). وهكذا، بحلول الوقت الذي خرج فيه من المتاهات الكربونية، كان لدى السلف المحتمل للثدييات حاسة شم متطورة، وحدبات سمعية في سقف الدماغ المتوسط، وبدائية قشرية تدمج مراكز الشم الجنسية والحركية والذوقية (الشكل 1). ثالثا-27، . أ،ب).
يطرح سؤال طبيعي تمامًا حول مصير هذه المخلوقات في المستقبل. يُفترض عادةً أن أسلاف الثدييات الصغيرة كانوا يشمون فرائسهم في أرض الغابة ليلاً، ويختبئون في الجحور أو بين جذور الأشجار أثناء النهار. وهذا افتراض عادل تماما، على الرغم من أنه يفسر فقط إمكانية تطوير حاسة الشم. ومع ذلك، مع نمط الحياة هذا، لا يمكن اكتشاف أي محفزات إضافية لتطوير القشرة المخية الحديثة، وخاصة نصفي الكرة المخيخية. على العكس من ذلك، تمتلك الحيوانات الشفقية أكثر من مجرد مخيخ متواضع. يتطلب التطور السريع للمراكز القشرية الحسية الحركية والمخيخ بيئة ثلاثية الأبعاد معقدة بشكل لا يصدق لم يسبق للفقاريات أن واجهتها من قبل. يجب أن نفترض أن التربة ليست هي التي تسببت في ظهور الحساسية الجسدية المتقدمة، بل بيئة أخرى.
في البحث عن بيئة تطور الثدييات، يمكن تقديم مساعدة كبيرة من خلال تحليل نظام مستقبلات آخر، وهو أمر يصعب المبالغة في تقديره - الحساسية الجسدية. اكتسب تكامل الثدييات مجموعة مذهلة من المستقبلات الميكانيكية من أنواع مختلفة. إنهم متخصصون في إدراك أنواع مختلفة من الاهتزازات والضغط واللمس والتدفئة والتبريد. لا يحتاج سكان التربة على الإطلاق إلى مثل هذه المجموعة المتنوعة من المستقبلات الجلدية، خاصة وأن ثدييات التربة الحديثة (فئران الخلد العارية) لديها شعر منخفض. من غير المحتمل أن يكون هناك نظام حسي جسدي متطور وشعر قد نشأ في الحيوانات التي تعيش أسلوب حياة شبه تحت الأرض.
على ما يبدو، فإن أسلاف الثدييات الزواحف، بعد أن تركوا الأنقاض الكربونية، انتقلوا إلى تيجان الأشجار (انظر الشكل III-27، الخامس،ز). تبدو "الهجرة" العمودية من مصدات الرياح النباتية ذات الإضاءة الضعيفة إلى عالم الشفق الخاص بتيجان الأشجار أمرًا طبيعيًا تمامًا. ولم يكن هذا التحول تغييرا جذريا في بيولوجيا أسلاف الثدييات من الزواحف. تم الحفاظ على بيئة معيشية ثلاثية الأبعاد مماثلة وأهمية الجهاز الدهليزي المتطور بالفعل. ومن المحتمل أن يكون الانتقال من المستويات الأدنى من حطام الغابات الكربونية إلى مظلة الأشجار قد حدث بشكل متكرر، ولكن بنتائج مختلفة. فقط بعد ظهور التخصص الأساسي لدماغ الزواحف وفقًا للنوع الشمي كانت المتطلبات الأساسية اللازمة لتشكيل المجموعة "الشجرية" من الثدييات القديمة قادرة على الظهور. في تيجان الأشجار الشفقية، من الضروري تحديد مجموعة من عمليات الاستحواذ الحسية العصبية والتحليلية والإنجابية المعروفة في الثدييات الحديثة.
إن حياة الكائنات الكبيرة في تيجان الأشجار تستبعد عمليا التكاثر في الأعشاش أو التجاويف. بالنسبة للحيوانات الصغيرة ذات حاسة الشم المتطورة، كانت مجموعة البيض الأخرى ولا تزال غذاءً مثاليًا ويمكن الوصول إليه، لذلك تم تطوير الحيوية الموروثة من أسلاف الزواحف. كان من الضروري إطالة نمو الجنين داخل الرحم قدر الإمكان. هذا جعل من الممكن تجنب تكوين العش والارتباط بمنطقة معينة. انتقلت الأم لتناول الطعام مع الشبل، مما زاد من احتمالية بقائهم على قيد الحياة.
إن أبسط طريقة لزيادة مدة التطور داخل الرحم هي رفض إطعام الجنين من صفار البيض. لا يمكن زيادة احتياطيات الصفار إلى أجل غير مسمى في رحم الأم. إنه أكثر فعالية بكثير استخدام تبادل انتشار بسيط للأكسجين والماء والأيضات بين جدار الكيس المحي والرحم. على ما يبدو، تم استخدام هذه الطريقة لحل مشكلة التطور داخل الرحم للثدييات القديمة. كان أسلاف الثدييات الشجرية حيوانات صغيرة جدًا. وقد سمح لهم ذلك بتنمية الأجنة إلى أحجام قابلة للحياة بالكامل باستخدام مشيمة الصفار. تستخدم الجرابيات الحديثة استراتيجية إنجابية مماثلة. ومع ذلك، فإن مشيمة صفارهم تسمح لهم بنمو جنين صغير فقط، والذي يجب نقله إلى كيس به الغدد الثديية. وبما أن الثدييات القديمة كانت صغيرة، فمن المحتمل أنه لم تكن هناك حاجة لتربية الأجنة في أكياس. فقط مع زيادة حجم الحيوانات يمكن أن تنشأ صعوبات في نمو الأجنة الكبيرة. حلت الحيوانات السفلية هذه المشكلة بمساعدة الكيس، والثدييات الأعلى - بمساعدة المشيمة (جيمسون، 1988).
جنبا إلى جنب مع تطوير استراتيجيات الإنجاب الفعالة في الثدييات القديمة، يجب أن يكون النظام الحسي الحركي قد مر بأكثر التغييرات وضوحا. في تيجان الأشجار، يكون الحمل على الجهاز الدهليزي أعلى عدة مرات حتى في بيئة مائية ثلاثية الأبعاد. حتى لو قامت السمكة بحركة خاطئة عند السباحة، فإن ذلك لا يؤدي إلى عواقب مميتة. يتم الحفاظ على الاعتماد على الماء في أي موقف ويسمح لك بتصحيح الأخطاء الحركية. بالنسبة للفقاريات المائية الأولية، تكون متطلبات النظام الحسي الحركي أقل أهمية بكثير من الحيوانات التي تعيش على أغصان الأشجار ولا تستطيع الطيران. يمكن أن يكون للأخطاء الحسية على فروع الأشجار عواقب وخيمة. أصبحت جاذبية الكوكب فاحصًا قاسيًا للزواحف التي انتقلت من أنقاض الكربون إلى الطبقة العليا من الغابة. كما فرضت حدًا لحجم جسم أسلاف الثدييات. ببساطة، لا تستطيع الحيوانات الكبيرة أن تنجو من أخطاء تطوير جهاز دهليزي ونظام حسي حركي مثالي. يؤدي سقوط الحيوانات الكبيرة من ارتفاع كبير دائمًا إلى الموت أو الضرر غير المعوض، وبالتالي فإن الحجم الخطي لأسلاف الثدييات لا يمكن أن يتجاوز عدة عشرات من السنتيمترات. كان على الحيوان الصغير والرشيق أن يكتسب بسرعة ليس فقط جهازًا دهليزيًا مثاليًا، بل طور أيضًا حساسية جسدية. يتم تمثيل هذا المركب الحسي على نطاق واسع في نصفي الكرة المخيخية والقشرة المخية الحديثة للثدييات.
من بين مستقبلات الجلد هناك مستقبلات تتكيف مع أنواع مختلفة من الاهتزازات. ظهرت أنظمة خاصة ذات أوقات تكيف مختلفة من أجل إدراك التقلبات. لن تكون مثل هذه المستقبلات الاهتزازية المتنوعة والمتخصصة في الجلد ضرورية على الإطلاق إذا كان أسلاف الفقاريات يبحثون عن فريسة على الأرض وفي فضلات الأوراق المتساقطة. على العكس من ذلك، فإن أغصان الأشجار وجذوعها تنقل أي اهتزازات بشكل مثالي. قد تحتوي هذه الاهتزازات على معلومات حول الفريسة، أو حيوان من الجنس الآخر، أو اقتراب حيوان مفترس خطير. كان لا بد من تمييز مثل هذه الإشارات عن الاهتزازات الآمنة ولكن المتنوعة للأشجار نفسها، وبالتالي فإن تطور الحساسية الجسدية في الزواحف الشجرية كان له ما يبرره بيولوجيًا تمامًا. في المرحلة الأولى من تطور أسلاف الثدييات الزواحف، ربما كانت حساسية المستقبلات الميكانيكية للجلد بعيدة كل البعد عن الكمال كما هو الحال في الحيوانات الحديثة. ويمكن تعويض هذا النقص من خلال تطوير تكوينات حساسة متخصصة. ومع ذلك، فإن المستقبلات المغلفة المعقدة مثل روفيني، باتشيني، مايسنر، أو قوارير طرفية لكراوس لا يمكن أن تنشأ على الفور لأداء وظائفها المتخصصة.
على ما يبدو، في المرحلة الأولى من تطور الحساسية الجسدية، تم استخدام النهايات العصبية الحرة، والتي تم تطويرها بشكل جيد في جميع الفقاريات. تكمن الصعوبة في أن النهايات العصبية الحرة لها قدرات حسية محدودة. إن الزيادة البسيطة في عددها في الأدمة لن تكون قادرة على حل المشاكل الحسية الجسدية المعقدة لأسلاف الثدييات الشجرية.
تم تحقيق زيادة الحساسية الميكانيكية الجسدية باستخدام الشعر (سبيرمان ورايلي، 1980). أصبح الشعر نوعًا من مكبرات الإشارة الميكانيكية. في الواقع، أسهل طريقة لتضخيم الإشارة الميكانيكية هي إنشاء رافعة أرخميدس غير متساوية التسليح. ستصبح الذراع الطويلة كاشفًا ميكانيكيًا، والذراع القصيرة ستصبح مستقبلًا مرتبطًا بنهاية عصبية حرة. من الواضح أن حساسية مثل هذا النظام سيتم تحديدها من خلال شكل وحجم وكتلة الرافعة وصلابتها وحساسية النهاية العصبية. إذا كان هناك العديد من هذه المستقبلات، فسيتم ضمان تمايز المعلومات الجسدية في الاتجاه والقوة والتردد. من الممكن أن يكون تطوير مثل هذا النظام المستقبلي الجسدي المتخصص قد أدى إلى ظهور الشعر المستقبلي (Hudspeth, 1985). تم استخدامه لاحقًا للحفاظ على الحرارة، مما أدى إلى إخفاء وظيفتها الأساسية. تتم الإشارة أيضًا إلى أصل مستقبل الشعر من خلال تطور نظامه العضلي. يمكن إجراء التنظيم الدقيق للتبادل الحراري بطرق فسيولوجية أخرى، ولكن لا توجد طريقة أخرى لتغيير حساسية المستقبلات الميكانيكية التي تربط بصيلات الشعر ديناميكيًا، لذلك في حالة الخطر، يقف شعر العديد من الحيوانات بشكل انعكاسي. وهذا يزيد من الحساسية الميكانيكية لخط الشعر نتيجة لتوتر "رافعة" المستقبل.
في الماضي البعيد، أدى التوتر في شعر المستقبلات لدى أسلاف الثدييات إلى زيادة دقة المعلومات الحسية الجسدية. هذا جعل من الممكن اختيار شكل مناسب من السلوك استجابة للموقف الذي نشأ. انطلاقا من الدعم العصبي، نشأت هذه الآلية لزيادة الحساسية الجسدية في فجر تطور الثدييات. لقد نجا حتى يومنا هذا كرد فعل لا إرادي لأي إثارة غير متوقعة. ونتيجة لذلك، تطورت الحساسية الجسدية الأولية لأسلاف الثدييات الزاحفة على أساس النهايات العصبية الحرة المرتبطة بالشعر المستقبلي. الدليل غير المباشر لصالح وجهة النظر هذه هو التعصيب العالي لأعمدة الشعر وبصيلات الشعر. في بعض الحيوانات، يمكن تجميع ما يصل إلى 20 من الألياف العصبية الحسية حول قاعدة الشعر. يتمتع نظام المستقبلات الميكانيكية هذا بأدنى عتبة إثارة وهو حساس للاهتزازات بتردد يبلغ حوالي 35 هرتز.
من خلال توفير زيادة في الحساسية الجسدية بالطريقة الأكثر بدائية، وضع أسلاف الثدييات الأساس للتطور طويل المدى للمستقبلات المغلفة المثالية. ولن تصبح أكثر فعالية من النهايات العصبية الحرة والنهايات المرتبطة بها إلا بعد ملايين السنين. كان المنتج الثانوي لتشكيل النظام الحسي الجسدي الأولي هو الشعر البدائي. يبدو أن تطورها الإضافي كطبقة عازلة للحرارة قد حدث في وقت لاحق بكثير من تكوين الوظائف الحسية الميكانيكية.
بالتوازي مع الجهاز الحسي المحيطي، تم تطوير آليات مركزية لتحليل الإشارات الجسدية واستقبال الحس العميق. إنها الحساسية الجسدية والجهاز الحركي الذي يتم تمثيله بمجالات واسعة في القشرة المخية الحديثة للثدييات المصابة بالدماغ (انظر الشكل III-24). ومن الواضح أن الحاجة إلى تطوير السيطرة القشرية على هذين النظامين أصبحت أحد الأسباب الرئيسية لتطور الدماغ الأمامي. يشار إلى ذلك من خلال التطور الموازي للعقد القاعدية (العقد القاعدية) للثدييات. مثل هذه الأورام المتخصصة الكبيرة في الجزء البطني من الدماغ الأمامي لم تنشأ من قبل في الفقاريات الأخرى (Reiner، Brauth، Karten، 1984). ومن المثير للاهتمام أن نلاحظ أن هذه المراكز النووية الضخمة تعالج المعلومات الحسية والحركية التي تأتي من أجزاء أخرى من الدماغ. إنها تعفي القشرة الحسية الحركية من التحكم في الحركات اللاإرادية.
وينبغي التأكيد على أنه بالتوازي مع التوسع في تمثيل المستقبلات الجسدية في القشرة المخية الحديثة، تم تشكيل اتصالات مماثلة لمستقبلات الجلد مع نصفي الكرة المخيخية. تم العثور على نصفي الكرة المخيخية المقترنة فقط في الثدييات بسبب تطور الحساسية الجسدية الهائلة وتنسيق الحركات المعقدة. لا يمكن ربط هذا التطور للمخيخ بأي ظروف قياسية في تاريخ الفقاريات قبل الثدييات. وحتى البيئة المائية ثلاثية الأبعاد التي تطورت فيها الفقاريات البدائية لمئات الملايين من السنين لم تتمكن من رفع أنظمتها الحسية الحركية إلى نفس مستوى التطور الذي حققته الثدييات.
تشكل المخيخ "الثديي" في ثلاثيات الأسنان خلال 30 إلى 40 مليون سنة فقط. وينبغي البحث عن سبب ظهوره في تيجان الأشجار العالية، حيث تعتمد حياة أي حيوان على فعالية تحليل الإشارات الجسدية وتنسيق حركات الجسم كله. في الثدييات، سطح المخيخ بأكمله مشغول بقشرة معقدة التنظيم، والتي تتكون من خلايا عصبية متخصصة. يتم تمثيل كل سطح مستقبلي من الجسم بمنطقة محددة بدقة من القشرة المخيخية. وأدى ذلك إلى زيادة مساحة سطح الهياكل القشرية لمخيخ الثدييات آلاف المرات مقارنة بالمخيخ الزاحف. في الواقع، نتيجة للتوسع الجانبي، ظهر نصفي الكرة المخيخية المقترنة. كانت نتيجة تطور الوصلات المخيخية بين نصفي الكرة الأرضية هي تكوين جسر الدماغ الخلفي للثدييات، وهو غير موجود في الزواحف والطيور. كان سبب تشكيل الجسر هو الحاجة إلى المقارنة التشغيلية المستمرة للمعلومات الجسدية القادمة من النصف الأيمن والأيسر من الجسم، والتصحيح الحركي لوضع الجسم. يعتمد بقاء الثدييات القديمة في تيجان الأشجار بشكل مباشر على تطور الجهاز التحليلي للحساسية الجسدية والحسية. أصبح المخيخ نوعًا من الآلات الحركية التي تدمج الإشارات الجسدية والحسية والدهليزية. ومن خلال أداء هذه الوظائف، سمحت لأسلاف الثدييات بحل مشاكل الحركة دون وعي في بيئة معقدة ثلاثية الأبعاد.
يساعد تطور الثدييات القديمة في مظلات الأشجار على تفسير التطور المحدد لأعضاء الحواس الأخرى وتمثيلها الدماغي. تتطلب البيئة المعقدة ثلاثية الأبعاد طرقًا جديدة تمامًا لتقييم المساحة المحيطة من الرؤية المبسطة لأسلاف الثدييات. كان من الضروري ليس فقط رؤية الكائن، ولكن تحديد المسافة إليه بدقة شديدة وتقييم خصائصه. عادة ما يؤدي سوء تقدير المسافة إلى فرع في مظلة الشجرة إلى خسارة الأرواح. إن الرؤية المجهرية والتمثيل القشري لهذا النظام في الدماغ الأمامي له ما يبرره تمامًا.
يجب التأكيد على أن الرؤية والحساسية الجسدية واستقبال الحس العميق والجهاز الدهليزي للأذن الداخلية هي المدخلات الحسية الرئيسية للنواة الدهليزية في جذع الدماغ. يتيح دمج هذه الإشارات للثدييات وضع أجسامها في الفضاء والتحكم في دقة تحركاتها. تعتبر النوى الدهليزية للثدييات تكوينًا فريدًا. وهي أكثر تطوراً بكثير من تلك الموجودة في الزواحف والطيور. من الواضح أن مثل هذا النظام متعدد الوظائف للتحكم الدهليزي والحركي لا يمكن أن يتطور إلا في ظل الظروف القاسية لمظلات الأشجار. في مثل هذه البيئة، كانت هناك جميع الظروف اللازمة لتشكيل نظام سمعي فريد من نوعه للثدييات. ربما تكون الأذن الخارجية، التي يمكن توجيهها نحو مصدر الصوت، قد نشأت في البيئة الصوتية المعقدة لمظلات الأشجار. تمتلك الثدييات الشجرية الحديثة مثل هذه الأصداف السمعية الخارجية. بعد أن اكتسبت السمات الهيكلية المدرجة للجهاز العصبي في تيجان الأشجار ، "نزلت" الثدييات مرارًا وتكرارًا إلى الأرض. كانت الكائنات الأحادية هي أول من عاد إلى الوجود غير الأرضي (انظر الشكل 111-27، ج-و)،ثم الجرابيات، وبعد ذلك كله، الثدييات المشيمية (انظر الشكل III-27، e-m). على ما يبدو، تم تشكيل الخفافيش والرئيسيات بالكامل في تيجان الأشجار. كان انتقال الرئيسيات إلى الوجود الأرضي هو الخطوة الأولى نحو ظهور الإنسان.
إن أهم ما اكتسبه دماغ الثدييات التي تعيش في تيجان الأشجار هو القدرة على التنبؤ بالأحداث. إن القدرة على التنبؤ بحدث ما، أو نتيجة الحركة، أو عواقب الصيد أو الصراع بين الأنواع، هي أيضًا ما يميز الثدييات الحديثة. إن قدرة الجهاز العصبي على التنبؤ بنتيجة الإجراء الذي لم يكتمل بعد كانت غائبة في الفقاريات الأخرى. لقد دفعت الثدييات ثمناً باهظاً لهذه القدرة من خلال الأخطاء التي ارتكبتها بعيداً عن الأرض. بعد أن نزلت إلى الأرض للمرة الثانية، لم تمتلك الثدييات مراكز ترابطية من نوع الزواحف فحسب، بل امتلكت أيضًا قدرة متواضعة على تقييم نتائج الإجراءات الفورية. يعتمد هذا الاكتساب الوظيفي لدى الثدييات على وفرة الخلايا العصبية والوصلات التي تشكلت في القشرة المخية الحديثة. فقط الذاكرة الزائدة والخبرة الفردية سمحت للثدييات باحتلال موقع مهيمن في عالم الحيوان.
نظرية الوسائط الانتقالية
يعتمد تطور الجهاز العصبي للفقاريات على الأنماط المورفولوجية العامة. إنها تتعلق بالتغيرات الكمية والنوعية في الجهاز العصبي المركزي والمحيطي. ومع ذلك، على عكس أجهزة الجسم الأخرى، فإن أي تغييرات هيكلية تؤدي إلى إعادة هيكلة عميقة للسلوك. والنتيجة هي تغيير في أشكال تفاعل الكائن الحي مع البيئة الخارجية. الخصائص الشكلية الجديدة للجهاز العصبي لا تؤدي دائمًا إلى نتائج إيجابية. أصبحت بعض هذه الخصائص أساسًا للازدهار قصير المدى لمجموعة أو تخصص مسدود، والبعض الآخر أعطى الفقاريات الفرصة لإتقان موارد لا نهاية لها وفتح مسارات واعدة للتطور. في التاريخ الطبيعي للجهاز العصبي، كانت ولا تزال هناك قرارات مورفولوجية تحكم على أصحابها بالانقراض الحتمي والازدهار. معظم الحيوانات الحديثة هي أمثلة على التكيفات الناجحة إلى حد ما، ولكن طريق مسدود. كان اختفائهم محددًا مسبقًا في لحظة بداية التخصص الهيكلي للجهاز العصبي.
يتمتع الجهاز العصبي بخاصية واحدة رائعة: فهو قادر على تغيير سلوك الحيوان والنشاط الفسيولوجي لأعضائه على الفور تقريبًا، ثم استعادة الوضع الأصلي بنفس السرعة. إن قابلية عكس التغييرات السريعة والضرورية للغاية تجعلها أداة لا تقدر بثمن في العالم البيولوجي. ومع ذلك، فإن نطاق إعادة الهيكلة المحتملة للجهاز العصبي محدود ببنيته. يمكن للدماغ أن يقدم فقط تلك المجموعة من القرارات الغريزية أو الترابطية التي توفرها الأنظمة الحسية الحركية للجسم. لن يلوح الدب بمخالبه، حتى لو كان يستطيع الطيران بالفعل. فقط الشخص المتحضر هو الذي يقرر بسهولة القيام بمثل هذه الإجراءات، لأن دماغه فقد الاتصال تقريبًا بالعالم الحقيقي للكوكب. بمعنى آخر، جميع الفقاريات هي أسرى الماضي التطوري لجهازهم العصبي. من خلال السماح للحيوانات بالتكيف بسرعة مع التغيرات اللحظية في البيئة، يشكل الدماغ نوعًا من الإطار الخفي لأقصى قدر ممكن من التغييرات. هذه الأطر هي التي تحدد مسبقًا حدود التغييرات التكيفية القابلة للعكس في سلوك نوع معين.
يعد التطور المورفولوجي للجهاز العصبي ضروريًا كأداة لتوسيع حدود القدرات التكيفية. تؤدي التغيرات الهيكلية في الدماغ إلى إزالة القيود من بعض ردود الفعل السلوكية وتشكيل ردود أفعال أخرى. يمكن أن تستمر هذه العملية حتى يصبح الدماغ متخصصًا جدًا بحيث لا يتمكن من إجراء المزيد من التغييرات. ومع ذلك، تظل التغييرات الكمية أو النوعية في الجهاز العصبي هي السبيل الوحيد لتجاوز القدرات القياسية. ويجب التأكيد على أن التغيرات الكمية في الجهاز العصبي يمكن أن تحدث بشكل أسرع بكثير من التغيرات النوعية. فهي المورد الأساسي للتكيفات الهيكلية للأنسجة العصبية. إن التغيرات المورفولوجية النوعية في الدماغ صعبة للغاية وتتطلب عادة ظروفًا خاصة أو فترة طويلة. يتم التوسط في هذا الاختلاف بين التغيرات الكمية والنوعية في الهياكل العصبية من خلال الموقع الخاص للدماغ والحبل الشوكي في نظام أعضاء وأنسجة الفقاريات.
لا يشارك الجهاز العصبي المركزي في التفاعلات الميكانيكية الحيوية للجسم مع البيئة الخارجية. هذا لا يعني أنه عندما تنقبض العضلات، أو تتحرك المفاصل، أو يتشوه الجلد، أو يتحرك الطعام عبر الأمعاء، فإن الأعصاب لا تتعرض لضغط ميكانيكي. لقد عرفوا القوة والليونة ويمكنهم تحمل الأحمال الصغيرة والقصيرة المدى. ومع ذلك، نحن لا نتحدث عن الخصائص الميكانيكية للجهاز العصبي. على العكس من ذلك، فإن الأمر الأكثر إثارة للاهتمام بالنسبة للتشكل التطوري هو أن الجهاز العصبي محمي للغاية من أي ضغوط، باستثناء المستقبلات الميكانيكية المتخصصة. يحدث التطور الكامل للدماغ والحبل الشوكي داخل الجمجمة وتحت حماية الأقواس العصبية للفقرات. ويتم فصلها عن العناصر الهيكلية بثلاثة سحايا والسائل النخاعي. ومع ذلك، فإن شكل الجهاز العصبي المركزي يختلف قليلاً باختلاف الهيكل العظمي. ويكفي أن نذكر أنه في الفترة الجنينية للنمو، يكون الدماغ والحبل الشوكي هما المحفزين للتمايز الهيكلي، وليس العكس. ومن العدل أن نقول إن شكل الجمجمة والأقواس العصبية للفقرات ثانوي بالنسبة لتشريح الجهاز العصبي المركزي. وبالتالي فإن الدماغ والحبل الشوكي يتغير شكلهما بغض النظر عن أي تحولات ميكانيكية حيوية في جسم الحيوان.
يحتوي هذا الوضع الخاص للدماغ على إمكانات هائلة لأي تحول هيكلي. المصدر الحقيقي للتغيرات الكمية هو التباين الفردي المستقر للجهاز العصبي. أظهرت الدراسات الخاصة التي أجريت على البرمائيات والزواحف والثدييات أنه في اليرقات القابلة للحياة أو الحيوانات الصغيرة التي تم الحصول عليها من زوج واحد من الوالدين، يكون التباين الكمي في الدماغ بنسبة 20-22٪ هو المعيار. تم حساب الخلايا العصبية في جميع أجزاء الدماغ وفي أجهزة التحليل الطرفية الرئيسية. تم العثور على بعض الاختلاف في قيم التباين اعتمادًا على منطقة الدماغ. تتميز الهياكل القديمة (الدماغ الخلفي والنخاع المستطيل) بتقلب بنسبة 7-13٪، وبالنسبة للهياكل الجديدة التطورية - 18-25٪. ومع ذلك، فإن التباين الكمي غطى جميع أجزاء الدماغ تقريبًا. تم تحديد حدود التباين المحددة في مجموعة متجانسة وراثيا من الحيوانات - أحفاد زوج واحد فقط من الوالدين.
إذا استخدمنا شخصًا لتقييم تباين الجهاز العصبي، فإن الاختلاف المزدوج في كتلة الدماغ لن يستنفد جميع المتغيرات الموجودة والقابلة للحياة (Savelyev، 1996)، وبالتالي فإن الاستنتاج الأكثر موضوعية سيكون حول التباين الطبيعي 15-25. ٪ تقلب الدماغ. وهذا يعني أن حيوانًا ما يختلف دائمًا عن الآخر بمقدار معين من الأنسجة العصبية. بالنسبة لفقدان الذاكرة، يمكن أن يتراوح هذا المورد من عدة آلاف إلى عشرات الملايين من الخلايا العصبية، وبالنسبة للحيوانات السلوية، من مئات الآلاف إلى عدة مليارات من الخلايا. وبالنظر إلى أن كل خلية عصبية لديها العديد من الاتصالات مع الخلايا الأخرى ويمكن أن تكون حاملة للذاكرة، يمكننا أن نفترض وجود اختلاف ملحوظ في سلوك الأفراد حتى في السكان الأكثر تجانسا. الأدلة الأخلاقية عديدة على تفرد السلوك وتغطي جميع مجموعات الفقاريات تقريبًا. وهذا يعني أنه يوجد في أي مجتمع أفراد يمكنهم حل مشكلات معينة بشكل أفضل أو أسوأ من الآخرين. إذا كان الوضع البيولوجي مستقرا، فلن يستفيد أحد من هذا الاختلاف في القدرات المحتملة للدماغ.
تصبح الاختلافات الكمية في الدماغ كبيرة عندما تكون البيئة غير مستقرة، أو عندما يكون هناك منافسة جنسية عالية، أو عندما يكون هناك مورد غذائي واضح ولكن لا يمكن الوصول إليه. لقد تبين أنها احتياطي حاسم عندما يتم استنفاد المجموعات الغريزية النقابية من أشكال السلوك الخاصة بالأنواع تمامًا. إذا كان الشكل الفردي للسلوك يوفر مكسبًا ملحوظًا في الوصول إلى الغذاء، فإنه يتم تعزيزه من خلال المزايا الإنجابية اللاحقة، وتزداد احتمالية الحفاظ على الخصائص الكمية لدماغ فرد معين. ومن الواضح أن هذه الآلية هي التي تكمن وراء التغيرات التكيفية واسعة النطاق في دماغ معظم الفقاريات الأولية. اعتمادًا على نوع التغذية وتطور الأعضاء الحسية، يزداد حجم دماغهم بشكل مختلف (انظر الفقرة 27). هذا المسار في تطور الجهاز العصبي فعال في حل مشاكل تكيفية معينة في إطار التصميم الحالي للجهاز العصبي. الأحداث التطورية الكبرى التي تؤدي إلى تغيير في الموائل وظهور أصناف منهجية جديدة ذات رتب عالية تتطلب تغييرات نوعية في الجهاز العصبي.
يتطلب ظهور هياكل جديدة نوعيًا في الجهاز العصبي وقتًا طويلاً وظروفًا خاصة جدًا. يجب أن تكون هذه الظروف مختلفة عن الموائل التقليدية ولها جاذبية لا تقاوم للفقاريات. ضمانات هذه الجاذبية هي الغذاء الوفير والتكاثر الناجح. إذا تم الحفاظ على هذه البيئة المفيدة بيولوجيا لفترة طويلة، فإن الحيوانات لديها فرصة للحصول على بنية عصبية جديدة نوعيا.
نشأت حالات بيئية قليلة من هذا القبيل في تاريخ الفقاريات، وتميزت جميعها بظهور حيوانات ذات هياكل جديدة نوعيًا للجهاز العصبي. الحدث الأول من هذا النوع كان ظهور الحبليات. كما هو موضح أعلاه، كان ظهور الحبليات حدثًا عشوائيًا إلى حد ما، وليس نمطًا تطوريًا قاتلًا (انظر الفقرة 26). استمرت مجموعة من الديدان المسطحة الصغيرة الشبيهة بالتوربيلا في العيش في المياه الضحلة الغنية بالغذاء. كونها مغذيات مرشحة وتعيش أسلوب حياة سلبي، حاولت هذه المخلوقات الشبيهة بالديدان المسطحة الحصول على موطئ قدم في المناطق الغذائية الأكثر ربحية. للقيام بذلك، قاموا بغمر الجزء الخلفي من أجسادهم في الرواسب السفلية. ينتشر هذا التثبيت على نطاق واسع بين اللافقاريات القاعية الحديثة. وكانت النتائج طويلة المدى لهذه الإجراءات التكيفية البسيطة للديدان القديمة هي الحبل العصبي الظهري والحبل الظهري العضلي، مما منع تشوهه. كان جوهر التغيرات النوعية في الجهاز العصبي المكون من سلسلتين أو أربع سلاسل لأسلاف الحبليات الشبيهة بالديدان هو عدة أحداث متتالية. وفي النسخة المزدوجة الجديلة، دارت الدودة بمقدار 90 درجة على أحد الأسطح الجانبية للجسم. مع بنية الجهاز العصبي المكونة من أربع سلاسل ، يُلاحظ اندماج السلاسل العصبية الظهرية والبطنية المقترنة. في كلتا الحالتين، بلغت إعادة الهيكلة النوعية للجهاز العصبي ذروتها في الاندماج المنقاري للعقد القطاعية لسلسلة العصب الظهري مع التكوين اللاحق للبطين المركزي. بالتوازي، كان هناك تقسيم لعقد السلسلة العصبية البطنية إلى مستوى العقد الجسدية (انظر الفقرة 26). لقد أصبحوا الأساس لتعصيب الأعضاء الداخلية. لم تكن Chordata لتظهر بدون بيئة انتقالية محددة. يضمن عمق المياه الضحلة ووفرة الغذاء والظروف المناسبة للتكاثر ازدهار أي مغذيات مرشحة سفلية. من بين الخيارات العديدة للتكيف مع مثل هذه البيئة المواتية، كان ظهور النمط الحبلي المورفي مجرد واحد من الخيارات الناجحة. وفي هذه الحالة، لعبت البيئة الغنية بالغذاء دورًا حاسمًا، الأمر الذي أصبح حافزًا للتغيرات المورفولوجية في العديد من الأنواع. حدث التطور الإضافي للحبليات في ظروف أكثر تنوعًا وأدى إلى ظهور التنوع الكامل للفقاريات المائية الأولية (انظر الفقرة 29).
خضع الدماغ للتغير النوعي الأساسي الثاني بعد وصول الفقاريات إلى الأرض. أدى هذا الحدث إلى تغييرات شكلية كبيرة في كل من الجهاز العصبي والأعضاء الأخرى. تم تشكيل الأطراف والتنفس الرئوي والأغطية المتخصصة وعدد من الخصائص الأخرى التي سمحت لرباعيات الأرجل القديمة بالانتقال إلى الوجود الأرضي. لا يمكن أن تحدث مثل هذه إعادة الترتيب المورفولوجي الوظيفية الواسعة لجهاز المحلل والجهاز المستجيب للجهاز العصبي في فترة قصيرة من الزمن وخارج بيئة انتقالية خاصة. كانت ضرورية بشكل خاص للتغيرات النوعية في الجهاز العصبي، لأنه من الناحية الكمية، من الواضح أن دماغ البرمائيات أدنى من الحيوانات المائية الأولية المتخصصة. عند الوصول إلى الأرض، ظهر الجهاز الشمي الميكعي الأنفي، والتحكم في التنفس، ومجموعة معقدة من مراكز التحكم في جذع الدماغ للأطراف في الجهاز العصبي للبرمائيات القديمة. خضعت الأنظمة البصرية والسمعية والدهليزية لتغييرات. يمكن أن يكون النظام البيئي الانتقالي بين الموائل المائية والبرية عبارة عن متاهات تربة غريبة أو حطام غابات الكربون (انظر الفقرة 31). في مثل هذه البيئة الانتقالية، يمكن استخدام حركات السباحة ودعم الزعانف لفترة طويلة. عندما كانت نسبة الرطوبة في المتاهات عالية، كان التنفس الجلدي والخياشيم وبراعم الرئة تعمل في وقت واحد. تم تبرير تطوير الأجهزة الحسية المائية والهواء والأنظمة الحركية في بيئة انتقالية من خلال المزايا البيولوجية التي يوفرها تطوير المناطق الغنية بالغذاء والمحمية جيدًا (انظر الفقرة 33). على ما يبدو، خلقت متاهات التربة وحطام الكربون من جذوع الأشجار بيئة انتقالية فريدة من نوعها للتطور التدريجي للجهاز العصبي للبرمائيات القديمة. فقط مع التطور طويل الأمد للتغيرات المورفولوجية يمكن أن تظهر مراكز العمود الفقري والنواة الحمراء للتحكم في الأطراف والعضو الميكعي الأنفي والبصلة الشمية الإضافية والمراكز السمعية والدهليزية الثانوية.
يمكن اعتبار الفترة التاريخية الثالثة في تطور الجهاز العصبي بمثابة تكوين دماغ الزواحف القديمة. كانت فترة الزواحف هي الأكثر إثمارًا في تاريخ الفقاريات. وضعت الزواحف المبادئ الأساسية للتطور الهيكلي للدماغ السلوي. وفي الزواحف تم تشكيل قسم ارتباطي لأول مرة في الجهاز العصبي. لقد نشأت من الدماغ المتوسط وكان اكتسابًا ناجحًا لدرجة أن الزواحف أصبحت المجموعة الأكثر هيمنة بين الفقاريات لملايين السنين. لم يكن من الممكن أن يتشكل مركز الدماغ المتوسط الترابطي بدون حاجة بيولوجية جدية. نشأت في بداية تطور الزواحف كوسيلة للتكيف مع البيئة العدوانية. احتاجت الزواحف القديمة إلى مقارنة المعلومات القادمة من حواس مختلفة باستمرار واتخاذ قرارات معقدة. كانت القرارات مدفوعة بالتكيف المستمر للسلوك مع الوضع المتغير بسرعة. لم تمتلك أدمغة الفقاريات والبرمائيات المائية الأولية هذه الخصائص. لقد اختاروا أحد أشكال السلوك الغريزية وفقًا لمبادئ مختلفة تمامًا. تم اختيار البرمائيات على أساس المنافسة بين مراكز الدماغ للتمثيل المحلل (الشكل III-28). وكانت المقارنة البسيطة لمستوى الإثارة شرطا كافيا لتنفيذ أحد البرامج الغريزية. كانت الزواحف أول من امتلك نوعًا جديدًا تمامًا من الأجهزة التحليلية (انظر الشكل III-28). لقد عملت على مبدأ مقارنة المعلومات الواردة من كل عضو حسي. بدأ الدور الحاسم يلعبه محتوى إشارة المحلل، وليس حقيقة الإثارة نفسها (انظر الفقرة 37). في واقع الأمر، طورت الزواحف أساسيات المبدأ الترابطي للبحث عن الحلول. ومن الواضح أننا نرى العلامات الأكثر بدائية لهذه الخاصية المدمرة للدماغ، لكنها نشأت على وجه التحديد في الزواحف. ربما كان تاريخ الزواحف أكثر ثراءً في التجارب العصبية مما يمكننا تخيله. يكفي أن نذكر اكتسابًا تاريخيًا آخر للزواحف - الهياكل القشرية للدماغ الأمامي (انظر الفقرة 39). قدمت المنافسة الجنسية، جنبًا إلى جنب مع التطور المذهل لحاسة الشم والجهاز الميكعي لدى الزواحف، الأساس لظهور الهياكل القشرية. تشكلت الهياكل القشرية للدماغ الأمامي على أساس مركز جديد يضمن تكامل الإشارات الجنسية مع بقية الحواس. تنافس هذا المركز التكاملي الجنسي لفترة قصيرة مع السقف الترابطي للدماغ المتوسط، لكن نشاطه لم يظهر إلا خلال موسم التكاثر. على ما يبدو، من أجل التكاثر الناجح، كان على الزواحف القديمة إخضاع جميع أجهزة الجسم لهذه المهمة، وكان لا بد من تجاهل أي أنشطة جانبية، بما في ذلك البحث عن الطعام (الشكل 1). الثالث-29).
لم يكن من الممكن ظهور الارتباطات والمراكز القشرية لدماغ الزواحف دون ظروف خاصة جدًا. ومع ذلك، لنفترض أن الزواحف القديمة استقرت ببساطة على سطح الأرض. وبدون منافسة جدية من البرمائيات والحشرات والنباتات، فإنها سرعان ما ستصبح المجموعة المهيمنة دون حدوث تغييرات عميقة في الجهاز العصبي. وفي مثل هذه الظروف، لا يمكن تقديم أي أساس حقيقي لتحسينه. علاوة على ذلك، من المستحيل العثور على أسباب خارجية لتشكيل حاسة الشم المتضخمة التي أدت إلى ظهور الهياكل القشرية للدماغ الأمامي. وبالتالي، تطورت الأحداث الحقيقية وفقًا لسيناريو مختلف تمامًا ولم يكن لها أي علاقة بالتشتت المثالي للزواحف عبر سطح الكوكب.
على الأرجح، من المرجح أن يكون هناك تطور طويل إلى حد ما للزواحف القديمة في بيئة انتقالية متخصصة. من الواضح أن هذا المكان البيئي لم يكن مناسبًا لتحقيق الازدهار الهادئ لمجموعة شابة من الفقاريات. على الأرجح، نشأت جميع عمليات الاستحواذ العصبية للزواحف كتكيفات تكيفية مع موطن معقد للغاية وبيئة تنافسية عدوانية. من الممكن أن تكون هذه الوسيلة عبارة عن حطام خشب الكربون من جذوع النباتات (انظر الفقرة 38). وقد استخدمت البرمائيات أيضًا هذه البيئة جزئيًا، لكن من الواضح أنها أتت إلى هناك للحصول على طعام وفير ومضمون. كان الطعام على الأرجح عبارة عن فقاريات مائية بدائية، والتي استخدمت حطام الكربون كأماكن مناسبة للتكاثر. مع مرور الوقت، قاموا بتغيير مواقع التكاثر أو انحسرت المياه. عندما يجف مصدر الغذاء لسبب أو لآخر، بدأ استخدام البرمائيات كغذاء لنوعها الخاص. أدى هذا إلى منافسة غير مسبوقة واختيار سريع للخصائص والقدرات الترابطية للدماغ.
وكانت البيئة الانتقالية لتكوين الزواحف عبارة عن حطام نبات الكربون، حيث وضعت البيئة ثلاثية الأبعاد متطلبات متزايدة على الجهاز الدهليزي والمحللات البعيدة. أدى غياب الضوء إلى رفع حاسة الشم إلى مستوى مختلف نوعيًا من التطور الشكلي الوظيفي. تم استخدامه كمحلل عن بعد ونظام مهم للتحكم في السلوك الجنسي. لقد تطور الجهاز السمعي بشكل نشط، وهو ليس أقل فعالية للتوجيه في الظلام.

على مدار عشرات الملايين من السنين من المنافسة الشرسة في متاهات نباتات الكربون، تطور دماغ الزواحف الفريد مع مجموعة مثالية إلى حد ما من الهياكل العصبية ومركز ارتباطي فعال. وبمساعدتها، تم حل مشاكل العثور على الطعام، والمنافسة، وتجنب الخطر، وما إلى ذلك. وعندما بدأ موسم التكاثر، أطاع الدماغ بأكمله البنية القشرية الجديدة في جدار الدماغ الأمامي. وأصبح مركزًا متخصصًا للتحكم في السلوك الجنسي، على عكس أي حيوان فقاري آخر قبل الزواحف. وهكذا، أصبح دماغ الزواحف القديمة النظام الأكثر مثالية لحل المهام البيولوجية الأكثر أهمية لأي نوع - البقاء والتكاثر. لكل مهمة، ظهر نظام تكاملي خاص بها، وهو قادر على إعادة توجيه كائن الزواحف بأكمله لحلها. مع مثل هذا المورد السلوكي، خرجت الزواحف من مهدها العدواني وسرعان ما أصبحت المجموعة المهيمنة على هذا الكوكب.
لا يمكن اعتبار ظهور دماغ الطيور حدثًا تطوريًا أساسيًا مرتبطًا بإعادة الهيكلة النوعية للدماغ. ربما كان من المفترض أن تختفي الطيور بعد وقت قصير من ظهورها. لقد كان تخصصًا تكيفيًا مسدودًا، والذي تم إنقاذه بفقدان حاسة الشم. لقد ورثت الطيور القديمة ركيزة عصبية ضخمة من الجهاز الشمي بسبب تغير تفضيلاتها الغذائية. بعد أن تحولوا إلى التغذية في حمامات ضحلة أو من الزعانف، توقفوا عن استخدام الرائحة كنظام تمييزي رئيسي. أصبحت الرؤية نظام التحليل الرئيسي، وأصبح السمع نظامًا إضافيًا (انظر الفقرة 43). عند الحصول على الطعام في الماء، تحركت الطيور القديمة على أطرافها الخلفية، مما أدى تدريجيا إلى انخفاض كبير في الحمل على الأطراف الأمامية والبدائية الجزئية لليد. وقد لعبت دور البيئة الانتقالية في هذه الحالة المياه الساحلية الضحلة الغنية بالغذاء، والتي ظلت جاذبة للطيور حتى يومنا هذا.
وعلى الرغم من أن التخصص الضيق للطيور يضمن انقراضها السريع، إلا أن الانتقال إلى السباحة والغوص بحثًا عن الطعام أدى إلى تطور أطراف أمامية تشبه الأجنحة. يبدو أن هذه المرحلة من تطور الطيور أنتجت طيور البطريق التي لم تطير أبدًا. أدى الغوص والسباحة باستخدام الأطراف الأمامية إلى خلق الظروف البدنية اللازمة لتطور العظام المجوفة والعضلات الصدرية القوية ونظام الكيس الهوائي للرئتين وغطاء الريش. على ما يبدو، أصبح الحصول على الطعام في المياه الباردة أحد الحوافز الرئيسية لاكتساب الدم الدافئ. تم استخدام أطراف السباحة التي تشبه الأجنحة ليس فقط للسباحة. استخدمت الطيور القديمة حركات رفرفة أطرافها الأمامية لنوع من "الجري على الماء"، والذي أصبح مرحلة انتقالية للطيران النشط (انظر الفقرة 44).

تم تشكيل الأجنحة وغطاء الريش للصيد في البيئة المائية، ولكن تم تكييفها واستخدامها للطيران. وفي هذه الحالة، أصبح الماء هو الوسيط الانتقالي. لقد خلقت جميع الظروف اللازمة للتراكم التدريجي للتغيرات في الجهاز العصبي للطيور، وبالتالي فإن ظهور الأجنحة والانتقال إلى الطيران لم يسبب تغييرات جذرية في الجهاز العصبي المركزي (انظر § 43). فيما يتعلق بانخفاض حاسة الشم لدى الطيور، تم تشكيل مراكز الارتباط على أساس الهياكل الأساسية للدماغ الأمامي. يتم تمثيل هذه المراكز من خلال النيو وفرط الحركة، والتي أصبحت الأساس لتشكيل سلوك الطيور المعقد والذاكرة وإضفاء الطابع الفردي على السلوك.
الثدييات هي مجموعة غريبة من الناحية العصبية. نشأت مزايا دماغهم على أساس تطوير الوظائف التكاملية للجهاز التناسلي. كما ذكر أعلاه، كان السبب الرئيسي لظهور الهياكل القشرية في دماغ الزواحف هو تطور العضو الميكعي الأنفي (جاكوبسون). تم تشكيل تمثيله المركزي خارج النوى الشمية القديمة للدماغ الأمامي. أصبحت المراكز الثانوية الرئيسية للشم الميكعي هي الهياكل القشرية المتواضعة للزواحف (انظر الفقرة 39). على هذه الركيزة المورفولوجية نشأت
تكامل السلوك الجنسي لكائن الزواحف بأكمله. جعلت هذه السيطرة المركزية من الممكن إخضاع الكائن الحي بأكمله لمهمة واحدة وتحقيق النجاح في التكاثر بشكل أكثر فعالية.
لقد ذهبت الثدييات إلى أبعد من الزواحف. في هذا الهيكل المورفولوجي الإنجابي التكاملي للدماغ الأمامي، تم تشكيل مركز ارتباطي من نوع جديد تمامًا. بدأت في أداء وظائف مراقبة تشغيل الأنظمة الحسية المنشأة بالفعل. ظلت آليات الدماغ المستقلة على مستوى المراكز القديمة، وجميع الوظائف المكتسبة المعقدة تطورت على مستوى قشرة الدماغ الأمامي. بالإضافة إلى حاسة الشم ومراكز التكامل الجنسي، يتميز دماغ الثدييات بتطور الجهاز الحسي الحركي وآليات التحكم الحركية. فقط في الثدييات شكل المخيخ نصفي الكرة الأرضية. لقد وصل إلى أبعاد هائلة لدرجة أن سطحه غالبًا ما يتجاوز حجم القشرة المخية الحديثة. علاوة على ذلك، فإن جزءًا كبيرًا، وأحيانًا كبيرًا، من القشرة المخية الحديثة نفسها يوفر وظائف جسدية وحسية وحركية.
لكي يظهر مثل هذا التخصص الغريب، هناك حاجة إلى بيئة أصلية للغاية. وكان حطام النبات الكربوني في حد ذاته بيئة معقدة ثلاثية الأبعاد للزواحف، لكن مخيخها لم يصل حتى إلى تطور مخيخ الطيور. لا بد أن البيئة الانتقالية لظهور الثدييات قد فرضت متطلبات عالية بشكل غير عادي على تحليل وضع الجسم وتنسيق الحركات. على سطح الأرض، فقط في فروع الأشجار يمكن أن تكون هناك مثل هذه المتطلبات الصارمة للتحكم الحركي. على ما يبدو، فإن جميع المزايا الحسية والحركية والشمية والسمعية الأساسية للثدييات تشكلت في مظلة الأشجار. يمكن لهذه البيئة الانتقالية أن تفسر ظهور القشرة المخية الحديثة وتطور الحساسية الجسدية، التي أصبحت أحد أعضاء الحواس الرئيسية (انظر الفقرة 48).
أدى تطور الحساسية الجسدية إلى تكوين مستقبلات الأدمة - الشعر. الشعر، الذي يعصبه النهايات العصبية الحرة، يزيد بشكل فعال من الحساسية الجسدية ومن ثم يتسبب في ظهور الشعر. الاستخدام المستمر للشعر للتنظيم الحراري يخفي غرضه الأساسي. في رؤوس الأشجار، ولأول مرة، نشأ متطلب جديد تمامًا للجهاز العصبي (انظر الفقرة 49). بالنسبة للثدييات الشجرية القديمة، لم يكن التحليل المقارن للمعلومات من مختلف الحواس كافيا. هذه الطريقة في عمل الأنظمة الترابطية لم تجعل من الممكن التنبؤ بالأحداث. في رؤوس الأشجار، أصبح التنبؤ بتطور الأحداث شرطًا حاسمًا للحصول على الغذاء والحفاظ على الحياة بشكل أساسي. الطيران وحده هو الذي يمكن أن ينقذ الثدييات من هذه المشاكل. إلا أن الخفافيش فقط هي التي لجأت إليها بعد تكوين المبادئ الأساسية لبنية دماغ الثدييات. تشمل العواقب الهيكلية الرئيسية للعيش في مظلات الأشجار القشرة المخية الحديثة، والمخيخ في نصف الكرة الغربي، والقدرة التنبؤية الضعيفة. هذه السمة التي تتمتع بها الثدييات، بعد هجرتها إلى التربة والبيئات المائية، خلقت لها مزايا سلوكية كبيرة. أصبحت القدرة على تقييم الأحداث المحتملة أداة للثدييات للسيطرة على الكوكب.
جميع التغييرات العميقة المدرجة في بنية الجهاز العصبي للفقاريات ناتجة عن تكيف الدماغ مع حياة الحيوانات في بيئة معينة. وبدون بيئة انتقالية طويلة المدى، لن يكون هناك وقت كافٍ للتغييرات في التنظيم الهيكلي للجهاز العصبي. إنه محافظ للغاية من الناحية النوعية والبلاستيكية من حيث الكمية بالنسبة للتحولات المورفولوجية السريعة والجذرية. إن افتراض وجود بيئات انتقالية قد يفسر أصول دماغ الفقاريات الحديثة.
من كتاب النملة، العائلة، المستعمرة مؤلف زاخاروف أناتولي ألكساندروفيتشظهور المستعمرة عادة ما يحدث تكوين الطبقات في وقت معين لكل نوع. هذه هي الطريقة التي تمت بها المرحلة الأولية لتحديد عش النمل الجديد في نملة الغابة المشعرة في عام 1967 في غابة شجرة التنوب بالقرب من موسكو (مصنع أخشاب سولنيشنوجورسك). خلال
من كتاب عشر أفكار عظيمة للعلوم. كيف يعمل عالمنا. المؤلف أتكينز بيترمقدمة ظهور فهم جاليليو أشار إلى نقطة تحول حيث اتخذت الجهود العلمية اتجاهًا جديدًا، حيث نهض العلماء - وهو مصطلح عفا عليه الزمن بالطبع في ذلك الوقت - من كراسيهم، وشككوا في صحة المحاولات السابقة
من كتاب الحلقة المفقودة بواسطة إيدي ميتلاندميتلاند إيدي الحلقة المفقودة (ظهور الإنسان - 2) الكتاب الثاني في سلسلة ظهور الإنسان يتحدث عن البحث عن الحقائق المتعلقة بالحلقة المفقودة في التطور المتعاقب للإنسان. وهي مخصصة لأسترالوبيثيكوس - حسب الاعتقاد السائد
من كتاب الأجنة والجينات والتطور بقلم راف رودولف أظهور الأجزاء يبدو تجزئة جنين ذبابة الفاكهة بشكل أساسي وكأنه سلسلة من الغزوات الجانبية للشريط الجرثومي، والتي تتشكل في وقت واحد تقريبًا في مرحلة المعدة. وعلى الرغم من الطبيعة الفسيفسائية الواضحة لهذه العملية، فإنه يمكن إثبات ذلك
من كتاب نظرية التغذية الكافية وعلم التغذية [الجداول في النص] مؤلف من كتاب نظرية التغذية الكافية وعلم التغذية [الجداول بالصور] مؤلف أوغوليف ألكسندر ميخائيلوفيتش9.3. ظهور الخلايا من المفترض أن تعتبر مرحلة نشوء الحياة على الأرض هي الفترة التي تشكلت فيها أبسط الأنظمة الخلوية، والتي أصبحت الخلية الأولية للحياة. يتم تناول المعلومات المتعلقة بهذه المشكلة في المراجعات المخصصة لها
من كتاب كيف نشأت الحياة وتطورت على الأرض مؤلف جريمياتسكي ميخائيل أنتونوفيتشالسادس. ظهور الحياة على الأرض من تجارب سبالانزاني وباستير، نعلم بالفعل أنه عند درجات الحرارة المرتفعة تتوقف الحياة. تموت معظم الكائنات الحية بالفعل عند درجة حرارة 70-80 درجة مئوية. وهذا يعني أن حياتهم تتطلب ظروف درجة حرارة معينة. مطلوب ل
من كتاب الدماغ والعقل والسلوك بقلم بلوم فلويد إيأجهزة تنظيم ضربات القلب في دماغ الثدييات - نوى فوق التصالبة في أواخر الستينيات، أجرى عالم الفسيولوجيا كورت ريختر سلسلة من التجارب على الفئران، في محاولة للعثور على مناطق الدماغ المسؤولة عن الإيقاع. لقد أدى إلى تدمير مناطق محددة من الدماغ – في أكثر من 200 مكان مختلف – في
من كتاب الحياة في أعماق العصور مؤلف تروفيموف بوريس الكسندروفيتشأصل الحياة: إن أصل الحياة، وجوهرها، هو من أصعب ألغاز العلم، فالحياة هي أعقد ظاهرة طبيعية معروفة لنا. لم ير أو يلاحظ أحد حدوثه. علاوة على ذلك، ليس بشكل مباشر أو غير مباشر
من كتاب أصل الدماغ مؤلف سافيليف سيرجي فياتشيسلافوفيتشالباب الثاني. ظهور الخلايا العصبية والدماغ كان سبب ظهور الجهاز العصبي هو انخفاض سرعة الحصول على المعلومات حول العالم الخارجي والداخلي للكائن الحي ذي التنظيم غير العصبي. تتكون أنسجتها من خلايا تحتوي على مواد كيميائية وكهرومغناطيسية مماثلة
من كتاب ولادة التعقيد [علم الأحياء التطوري اليوم: اكتشافات غير متوقعة وأسئلة جديدة] مؤلف ماركوف الكسندر فلاديميروفيتش§ 28. ظهور أجزاء من الدماغ الفترة المبكرة من تاريخ نشأة أسلاف الفقاريات، قبل تكوين هيكل عظمي جيد التنظيم، غامضة إلى حد ما. إذا افترضنا أن أشكال أسلاف الحبليات كانت مخلوقات رخوة يبلغ طولها حوالي 10-15 سم،
من كتاب الحالة الراهنة للمحيط الحيوي والسياسة البيئية المؤلف كوليسنيك يو.ظهور التعديلات ووراثتها باستخدام مثال ميتازون هناك مسألة نظرية مهمة أخرى يمكن أخذها في الاعتبار باستخدام مثالنا ميتازون وهي مسألة ظهور التعديلات التكيفية. هذا هو ما تسمى قدرة الجسم بشكل أو بآخر (أي،
من كتاب الأنثروبولوجيا ومفاهيم علم الأحياء مؤلف كورشانوف نيكولاي أناتوليفيتش4.2. نشوء التنظيم الخلوي أثبت العلماء أنه بعد تبخر الماء من حجم التفاعل، تتشكل تجمعات بلورية سائلة في جزيئات شحمية برمائية تشبه الدهون، حيث تترتب الجزيئات في طبقات دورية، كما في
من كتاب المؤلفظهور جنس هومو نظرًا لأن تطور البشر حدث بشكل غير متساوٍ وكان له طابع "فسيفسائي" ، فإن الحدود بين الممثلين القدامى لجنس هومو والأسترالوبيثيسينات غير واضحة للغاية. المعايير الشرطية للانتساب إلى جنس هومو هي حجم الدماغ (على الأقل
من كتاب المؤلفظهور وتطور الإنسان الحديث أصل الإنسان الحديث هو اللغز الأكثر إثارة للاهتمام في تكوين الإنسان. في الأنثروبولوجيا، تلقت جميع أشكال الإنسان الحديث الاسم العملي "العاقل". وكان معظم ممثليهم
من كتاب المؤلفملامح تطور دماغ الثدييات والبشر لا تزال المعدلات المرتفعة بشكل استثنائي لتطور دماغ الثدييات، وخاصة البشر، ليس لها تفسير واحد. علاوة على ذلك، عادة ما تتميز ناقلات العقول الكبيرة بأحجام الجسم الكبيرة، كبيرة
تطور الدماغ في الثدييات
يتميز تطور الدماغ بالزيادة التدريجية في كتلته وحجمه وتعقيد تنظيم القشرة المخية الحديثة. أثناء التطور، يتم دفع القشرة الأثرية والقشرة القديمة نحو الحاجز الدماغي.
في الحيوانات ذات الرتب المختلفة، يمكن ملاحظة التكوين التدريجي للطي.
تتميز الثدييات بعدة أنواع من تنظيم الدماغ.
نوع ليس الدماغإن تنظيم الدماغ هو سمة من سمات أحاديات المسلك (خلد الماء). مع هذا النوع من التنظيم، يتم التعبير باستمرار عن التلم المركزي، الذي يقسم الهياكل القشرية إلى الأجزاء الأمامية والجدارية. تهيمن القشرة القديمة من الناحية الكمية، بينما تحتل القشرة المخية الحديثة مساحة أصغر نسبيًا من الهياكل القشرية. السمة المميزة لهذا النوع من التنظيم هو غياب الجسم الثفني. في الدماغ عديم الدماغ، هناك تداخل بين المجالات القشرية لإسقاطات أنواع مختلفة من المشاعر. تتداخل الخلايا العصبية وتستقبل المعلومات من قنوات مختلفة. تتداخل القشرة مع المناطق الحركية والحسية. في الدماغ البيني، يتم تعميق التمايز بين النوى المهادية: لوحظ أن المجموعة تشكل في الحشرات مجموعة ترابطية من النوى. توجد في القشرة منطقة من العصبونات الحركية ألفا التي تنظم العصبونات الحركية ألفا في النخاع الشوكي، وهذا يؤدي إلى تكوين المخيخ الجديد في المخيخ، والذي يشارك في تنظيم النشاط الحركي.
تحتفظ الجرابيات بنفس تنظيم الدماغ.
نوع جيرينسيفاليتنظيم الدماغ هو سمة من سمات الحشرات والقوارض. ومع هذا النوع من التنظيم يظهر الجسم الثفني وتزداد مساحة القشرة الجديدة وتظهر الأخاديد والتلافيف.
في الحشرات، يتم تشكيل منطقتين متعددتي الحواس - نظائرها من المناطق الترابطية.
في القوارض، تتميز المناطق الأمامية والجدارية شكليا، لكنها لا تعمل بعد كمناطق ارتباط. يتم تنفيذ هذه الوظيفة من خلال عدة مجالات من القشرة المخية الحديثة ذات توطين مختلف. يوجد أيضًا في هذه المجموعة من الحيوانات تخصص واضح للمناطق الحسية لأنواع معينة من المشاعر، أي. لم تعد تتداخل مع بعضها البعض. في القوارض، يظهر خيار آخر لإسقاط المعلومات المرئية (في البداية، كان المفتاح الرئيسي هو الشبكية التكتلية (أحادية المسلك، الجرابيات، آكلات الحشرات)) - المهادية الشبكية. ويختلف من حيث أن الاختيار الرئيسي للمعلومات ومعالجتها يتم بواسطة الأجسام الركبية للمهاد. في القوارض، لوحظت الاتصالات المهادية القشرية التي تم تشكيلها بالفعل، ومع ذلك، فإن تمايزها الوظيفي ضعيف نسبيا.
نوع ديتروهيرينسيفاليكتنظيم الدماغ هو سمة من سمات الثدييات آكلة اللحوم. في الحيوانات المفترسة، بالإضافة إلى الأخاديد والتلافيف الثابتة، هناك تطور كبير في طي القشرة المخية الحديثة. يتم تمييز المناطق المتمايزة بوضوح: مناطق الإسقاط الخاصة (معلومات العملية من النظام الحسي)، والمناطق الحركية (الحركية)، والمناطق الترابطية جيدة التكوين، والتي يتم تمثيلها في الفص الجداري والفص الجبهي. المناطق الترابطية القشرية هي جزء من نظامين ترابطيين مهاديين قشريين:
- الجهاز المهادي الجداري مسؤول عن مكونات الجهاز العصبي العلوي: فهو يشارك في تكوين ردود الفعل الاتجاهية، ويشكل فكرة التخطيط المكاني للجسم، ويشارك في آليات الذاكرة قصيرة المدى.
- يرتبط النظام المهادي الجبهي بهياكل الجهاز الحوفي. يشكل متقبل الفعل (يقارن على مستوى الأشكال القشرية إلى أي مدى تتوافق النتيجة مع التكيف).
النوع البشرييتميز تنظيم الدماغ في الرئيسيات العليا بقشرة مخية حديثة مكونة من ست طبقات. جميع المناطق الوظيفية للقشرة لا تتداخل مع بعضها البعض، فهي تشكل ثلاثة أنظمة تكاملية:
- المهاد الجداري، ويشمل الحقول 5 و 7 من القشرة الترابطية ويؤدي الوظائف التالية: إثارة رد فعل توجيهي، وتكوين ذاكرة قصيرة المدى، والشعور بالتخطيط المكاني للجسم.
- يوفر نظام المهاد الأمامي تقييمًا للموقف: متقبل نتيجة الإجراء، يوفر آلية للذاكرة طويلة المدى، وتشكيل اتصالات مؤقتة دقيقة نتيجة للإجراءات السلوكية (أشكال أعلى من التثبيط).
- يوفر النظام المتكامل المهادي الصدغي تنظيمًا أعلى للوظائف الخضرية من القشرة من خلال المراكز التكاملية للجهاز العصبي المركزي، وفي المقام الأول منطقة ما تحت المهاد، ومجمع اللوزة، والعقد القاعدية.
فهرس:
1. نوزدراتشيف م. بدايات علم وظائف الأعضاء / أ.د.نوزدراتشيف، يو.آي.بازينوف، آي.أ.بارانيكوف وآخرون - سانت بطرسبرغ: دار لان للنشر، 2002، -1088 ص.
2. شميدت نيلسون ك. فسيولوجيا الحيوانات: التكيف والبيئة. الكتاب 1.- م: مير، 1982، -416 ص.
3. علم وظائف الأعضاء التطوري. الجزء 1 / إد. أكاد. إي إم كريبس. في السلسلة: "دليل علم وظائف الأعضاء" - ل: "العلم"، 1979، -603 ص.
يمتلك دماغ الثدييات من مختلف الرتب مراحل مختلفة من تمايز البنية الداخلية، مما يعكس مراحل تطور النشاط العصبي العالي الذي مرت به الثدييات في تطورها التاريخي.
في المذرق، الدماغ من
يتميز فقط بالمرحلة الأولية لتطور القبو النخاعي الثانوي - حيث يحتل النيباليوم جزءًا صغيرًا فقط من سقف الدماغ الأمامي. في الجرابيات، يشكل القسم الشمي من الدماغ الأمامي أيضًا الأجزاء السفلية والجانبية من نصفي الكرة الأرضية، ويتم تمثيل السقف فقط بواسطة قبو جديد - النيوباليوم. المرحلة التالية هي دماغ الحشرات والخفافيش، حيث تشكل القشرة الرمادية لنصفي الكرة الأرضية بالفعل قبوًا دماغيًا كاملاً، ويتم نقل القبو الأولي (الأرشيباليوم) إلى السطح الإنسي لنصفي الكرة الأرضية في شكل الحصين. تحتوي صوار النيباليوم على الشكل البدائي لحلقة من المسارات: الجسم الثفني (الجسم الثفني) والقبو (القبو). في معظم الثدييات المشيمية، تلعب المستقبلات الشمية، كما ذكرنا أعلاه، دورًا رائدًا في بيولوجيتها، على عكس الطيور، حيث تلعب الرؤية دورًا رائدًا. ولذلك، فإن مراكز الشم في الدماغ، والتي تتركز في الفصوص الشمية وفي الجزء السفلي من الدماغ الأمامي (النظام المخطط)، متطورة للغاية.كما تصل الإدراكات البصرية والسمعية إلى درجة عالية من التطور، وتتركز مراكزها في الدماغ المتوسط في المنطقة الرباعية التوائم. ولكن أبعد من ذلك، تتطور مراكز الدماغ العليا الثانوية للرؤية والسمع (الفصوص القذالية والصدغية لنصفي الكرة الأرضية) في القشرة الرمادية لنصفي الكرة الأرضية. تصل الوظيفة الحركية للأطراف إلى درجة عالية من التطور في الثدييات، وكذلك في الطيور، لذلك يتم عزل المراكز الحركية الثانوية الخاصة في القشرة الدماغية.
ترتبط هذه المراكز الترابطية الثانوية بالتطور القوي في دماغ الثدييات لأقسام جديدة من المخيخ: نصفي الكرة المخيخية، المتصلين بواسطة الصوار المستعرض للجسر.
تطور الدماغ بشكل مستقل في عدد من الثدييات - من الحشرات إلى الرئيسيات والبشر. في هذه السلسلة، من خلال نصف الليمور إلى الليمور والرئيسيات، لوحظ ضعف مراكز الشم، لأنه في جميع الأشكال الشجرية، تكتسب الرؤية والسمع المجسمة أهمية رائدة في علم الأحياء. ميزة أخرى مهمة لتحول دماغ الثدييات هي اكتساب القشرة الدماغية للطي على شكل أخاديد وفصيصات القشرة الرمادية. الثدييات السفلية - الحشرات والقوارض والأرنبيات والخفافيش - لها دماغ سلس. في الحيوانات العليا، يظهر نظام من الأخاديد الدماغية على نصفي الكرة الأمامية وعلى المخيخ. يصل الطي (الفصوص والأخاديد) إلى أقصى تطور له في الدماغ البشري، حيث تتركز جميع المراكز الترابطية الثانوية للنشاط العصبي العالي في القشرة الدماغية. يتطور الطي القشري أيضًا بسرعة في الثدييات المائية (زعنفيات الأقدام والحيتانيات) فيما يتعلق بالوظائف الجديدة لنشاط الدماغ في الماء.
الجميع لديه الجمجمةفي الحيوانات، يتحول الطرف الأمامي لأنبوب الدماغ إلى دماغ. في المراحل المبكرة من التطور الجنيني، الدماغ لامبريممثلة بثلاثة سماكات لجدران الطرف الأمامي لأنبوب الدماغ، والتي تتطور منها ثلاثة أجزاء من الدماغ لاحقًا: الأمامي والوسطى والخلفي أو المعيني. يرتبط الدماغ الأمامي - الدماغ المُقدم - بالعضو الشمي، وبالتالي فهو الدماغ الشمي؛ الدماغ المتوسط - الدماغ المتوسط - يرتبط بجهاز الرؤية، وبالتالي، هو الدماغ البصري وظيفيًا، وأخيرًا، الدماغ المؤخر، أو الدماغ المعيني - الدماغ المعيني - يرتبط بتطور الأعضاء VlIl وX أزواج من الأعصاب القحفية، أي مع أعضاء التوازن وعضو الخط الجانبي، وكذلك مع الأعضاء الداخلية، ونتيجة لذلك فهو أعلى مركز حركي، وينسق الجهاز الحركي بأكمله من خلال الحبل الشوكي ويتحكم في عمل الأعضاء الداخلية. في جميع حيوانات الجمجمة العليا الأخرى، يتم تمثيل الدماغ الجنيني بامتداد على شكل المثانة للأنبوب العصبي الموجود أمام الحبل الظهري. تسمى هذه الحويصلة ما قبل الحبلية بالحويصلة الدماغية الأولية - الدماغ القوسي (الشكل 156-7). الطية البطنية - plica ventralis (13) - سرعان ما تنفصل عن المثانة الدماغية الثانوية - ديترينفالون (12)، وتقع على شكل إبيكوردالي (فوق الحبل الظهري).

بالتزامن مع ظهور الطية البطنية، بسبب الانحناء البطني للدماغ المتوسط قبل الحبلي، ينفصل الدماغ المتوسط الثالث، أو الدماغ المتوسط، عن الدماغ المتوسط الثانوي (الشكل 157، نظام تحديد المواقع العالمي)؛ تُسمى الحويصلة أمام الحبل الدماغي بالدماغ الأمامي - الدماغ المُقدم (pc)، والحويصلة فوق الحبلية - الدماغ المُعين - الدماغ المُعين (rh). من السطح الظهري، يتم تحديد الأجزاء الثلاثة للدماغ بشكل حاد تمامًا عن بعضها البعض عن طريق الصوار المستعرض أو صوار الألياف العصبية، سواء أمام الدماغ المتوسط أو خلفه.
بعد ذلك، من الدماغ الأمامي على جانبي الصفيحة النهائية (بدلاً من المسام العصبية) ينشأ على شكل حويصلة دماغية مقترنة - الدماغ الانتهائي (B-9)، والجزء المتبقي من الدماغ الأمامي يشكل الدماغ البيني (10). ). في وقت واحد تقريبًا، يتم إطلاق الدماغ المؤخر (الدماغ التالي) من الدماغ المعين، الذي يتطور منه المخيخ لاحقًا، وفي الثدييات، بالإضافة إلى ذلك، تشكل الكتلة الرئيسية للدماغ المعيني النخاع المستطيل (الدماغ النخاعي)، أو النخاع المستطيل.
تسمى تجاويف الحويصلات الدماغية الثلاثة الأولية بالبطينات الدماغية. يتم فصل البطينين الدماغيين الجانبيين عن تجويف الدماغ الانتهائي (الشكل 158-I و II)؛ يسمى تجويف الدماغ البيني بالبطين الدماغي الثالث (III). يتواصل مع الجوانب الجانبية من خلال الثقب بين البطينين أو مونرو (8). نتيجة لنمو جدران المثانة الدماغية، يتحول بطين الدماغ المتوسط إلى قناة قناة الدماغ (7)؛ وهو يربط البطين الدماغي الثالث بالبطين الدماغي الرابع (IV)، ويقع الأخير في الدماغ المعيني، بين النخاع المستطيل والمخيخ، ويتواصل ذيليًا مع القناة المركزية للحبل الشوكي.
يتكون دماغ الحيوان البالغ من حويصلات الدماغ الموصوفة؛ في الوقت نفسه، تنمو الأقسام الفردية منها بشكل كبير للغاية، أو على العكس من ذلك، تحتفظ بحالتها الجنينية مدى الحياة. إن العامل المباشر الذي يحدد تطور الدماغ هو تمايز الأعضاء الحسية، خاصة مثل أعضاء الشم والبصر والسمع (في الأرضية)، وأعضاء الحركة الإرادية، وهو ما يصاحبه ظهور مراكز التنسيق المختلفة مع مساراتهم. وهكذا، في السلاخيين، الذين يتنقلون في حركاتهم بشكل رئيسي بمساعدة الرائحة، تصل الفصوص الشمية للدماغ الانتهائي إلى أقصى قدر من التطور في الدماغ. على العكس من ذلك، فإن أجهزة الرؤية في الأسماك العظمية لها أهمية كبيرة، ووفقًا لهذا، تتطور المراكز البصرية في الدماغ المتوسط بقوة أكبر، بينما يصبح الدماغ الشمي أقل تعقيدًا. يكون الدماغ المؤخر (المخيخ) أكثر اكتمالاً في الحيوانات سريعة الحركة، ولكنه في الحيوانات بطيئة الحركة يكون أوليًا تمامًا أو حتى غائبًا.
في الحيوانات الأرضية، يصبح الدماغ أكثر تعقيدًا في بنيته بسبب ظهور مراكز جديدة (عليا) في الدماغ الانتهائي وارتباطاتها مع أجزاء مختلفة من الدماغ والحبل الشوكي، ومع جميع أعضاء الحواس (الخارجية والداخلية) . ونتيجة لذلك يتطور قسم يسمى القشرة الدماغية في الدماغ الانتهائي، وهو المركز الأعلى الذي ينسق ويتحكم في جميع أنشطة الكائن الحيواني في البيئة الخارجية وجميع العمليات التي تحدث داخله، حيث أن جميع ردود الفعل الناشئة تكون مغلقة في الدماغ. القشرة الدماغية.
الدماغ الماسي
يتكون الدماغ المعين من النخاع المستطيل والدماغ المؤخر، ويمثلهما المخيخ والجسر.
النخاع
النخاع المستطيل س. الدماغ النخاعي (الشكل 159 و160-5) - يتم بناؤه بشكل موحد في الحيوانات، حيث تنطلق منه جميع الأعصاب القحفية، والتي ترتبط مباشرة بجهاز الخياشيم ومشتقاته، وجهاز التوازن والسمع، والأحشاء والقلب. وهو استمرار مباشر نحو رأس الحبل الشوكي، ولكنه يختلف بشكل حاد عن الأخير في بنيته. تم تشكيل النخاع المستطيل نتيجة لنمو الجدران الجانبية لأنبوب الدماغ الجنيني. نظرًا لأن الصفائح الجانبية ابتعدت عن بعضها البعض وكانت موجودة بشكل جانبي من الصفائح الرئيسية، فقد توسع تجويف الدماغ المعيني بشكل كبير وتحول إلى البطين الدماغي الرابع. يأخذ الجزء السفلي من البطين في الحيوانات العليا شكل حفرة على شكل ماسة، على سطحها، حتى في الثدييات البالغة، تكون الحدود بين الصفائح الجانبية والرئيسية مرئية بوضوح في شكل أخدود حدودي - التلم الحدودي . تعتبر الصفائح الجانبية ذات أهمية خاصة في الأسماك؛ إنهم يشكلون الفصوص السمعية الجانبية - الفص السمعي الجانبي - والتي يتم فصلها قحفيًا حتى في آذان الدماغ المعيني - الأذنيات. أنها تحتوي على نواة الزوج الثامن من الأعصاب، والتي ترتبط بجهاز التوازن، مع كامل عضلات الجسم (عبر الحبل الشوكي) ومع المخيخ. ومن بينها، تستحق نواة ديتر اهتمامًا خاصًا باعتبارها الأكثر ثباتًا في الحيوانات (الشكل 163-1).
نظرًا لتباعد الحواف الظهرية للصفائح الجانبية، فإن الصفيحة السقيفية ممتدة بشكل كبير، ولكن حتى في الفقاريات العليا البالغة تظل في الحالة الجنينية على شكل صفيحة ظهارية - الصفيحة الظهارية - تتكون من طبقة واحدة من البطانة العصبية الخلايا. يندمج مع أوعية الأم الحنون ويسمى في الحيوانات البالغة بالرق الذيلي النخاعي، أو الضفيرة المشيمية للبطين الدماغي الرابع - النخاع النخاعي الذيلي، resp. الضفيرة المشيمية البطينية الرباعية (الشكل 157-ب، 4). يحتوي على فتحات يتواصل من خلالها تجويف البطين مع الفضاء داخل القراب.
يفسر انفصال الصفائح الجانبية أيضًا عن الخروج الغريب للأعصاب من النخاع المستطيل: حيث توجد الأعصاب الحركية بشكل أكثر وسطيًا، وتقع الأعصاب الحسية بشكل جانبي أكثر. نظرًا لأن النخاع المستطيل يقع بين الحبل الشوكي والأجزاء الأمامية من المخ، التي يتكون منها المخ، والتي يفصل عنها برزخ ضيق (2)، فإن المسارات المركزية التي تسير في اتجاه أو آخر تقسم النخاع الرمادي إلى عدد من مجموعات النوى الفردية للخلايا العصبية. بعض النوى هي المراكز الخلوية للأعصاب القحفية من الخامس إلى الحادي عشر، وتمتد من النخاع المستطيل. على العكس من ذلك، يعمل البعض الآخر كمراكز وسيطة للمسارات المتجهة إلى الدماغ من الحبل الشوكي أو العكس. وتشمل هذه نواة الحويصلات الظهرية وأقدم مركز حركي - النواة الحركية للغطاء، الموجودة بالفعل في الأسماك. تتبع الألياف من نواة الحويصلة الظهرية في الحيوانات المائية إلى المخيخ والدماغ المتوسط، وفي الحيوانات الأرضية، بالإضافة إلى الدماغ البيني.
بفضل خلايا النواة الحركية للغطاء، يتم فصل ما يسمى بالزيتون في الحيوانات الأرضية: الأنف (العلوي) والذيلية (السفلية)، لتصبح مراكز الارتباط.
تظهر الزيتون الأنفي لأول مرة في البرمائيات بسبب تمايز القوقعة (جهاز السمع). يتم إدخالها في الفلم الجانبي، أي في حلقة من المسارات المتقاطعة التي تربط نواة العصب القوقعي (الزوج الثامن) بالمهاد البصري. توجد حزمة من هذه الألياف بشكل سطحي في الطرف الأمامي من النخاع المستطيل. تم تطويره بقوة أكبر في الثدييات ويعرف باسم الجسم شبه المنحرف - الجسم شبه المنحرف (الشكل 175-14).
الزيتون الذيلي محدد جيدًا في الطيور وخاصة في الثدييات؛ لديهم شكل حدوة حصان مطوي غريب ويستقبلون نبضات من نوى الحبل الظهري ومن الحبل الشوكي ومن المهاد البصري ويرسلون نبضات مرة أخرى إلى الحبل الشوكي وإلى المخيخ (الشكل 163).
تشكل المسارات الحركية التي تنشأ من القشرة الدماغية حزمًا في الثدييات على النخاع المستطيل البطني، المعروف باسم الأهرامات. يصلون إلى الحد الأقصى لحجمهم عند البشر.
الدماغ المؤخر
في الحيوانات السفلية، يتم تمثيل الدماغ المؤخر فقط من خلال المخيخ (المخيخ) (الشكل 159 و160-4). وظيفة المخيخ هي الحفاظ على التوازن وقوة العضلات، بالإضافة إلى تنسيق عمل العضلات أثناء الحركة، لذا فمن الطبيعي أن يصل إلى أقصى تطور له في الحيوانات التي تتحرك بسرعة (السباحة السريعة أو الطيران أو الجري) أو التحرك على طرفين. وهذا يحدد أيضًا التغيرات المرتبطة بالعمر. على سبيل المثال، في الرئيسيات العليا، يكون المخيخ صغيرًا عند الولادة ويشكل فقط 1/16-1/18 من المخ بأكمله، بينما يصل في البالغين إلى 1/8-1/9 من وزن الدماغ. ويصل الأمر إلى قدر أكبر من التعقيد من حيث التفاصيل الهيكلية في الحيوانات القادرة على القيام بحركات أكثر تمايزاً، أي في الثدييات.
المخيخ في الشكل البدائي عبارة عن جسم غير مزدوج على شكل صفيحة قصيرة ولكن واسعة، مبنية بالفعل في جلكيات من مادة الدماغ الرمادية والبيضاء. وبما أن وظيفة المخيخ تصبح أكثر تعقيدا في عدد من الحيوانات، فإن الصفيحة العصبية تنمو من الأمام إلى الخلف وتشكل في الوقت نفسه طيات عرضية أو في كلا الاتجاهين، أي إلى الأمام والخلف، كما هو الحال في أسماك القرش، أو إلى الخلف فقط كما هو الحال في الأسماك العظمية. في هذه الحالة، يتم فصل الجزء الأوسط - جسم المخيخ - الجسم المخيخي - والفصوص الجانبية لأذنيه - الأذنية - المرتبطة بجهاز التوازن. يتم أيضًا الحفاظ على الأجزاء الجانبية من المخيخ في الحيوانات الأرضية تحت اسم flocculus، والتي، مع ذلك، تم تطويرها بشكل سيء للغاية بسبب انخفاض أعضاء الخط الجانبي للحيوانات المائية. في الحيوانات الأرضية، يمثل المخيخ في شكله الأكثر بدائية أيضًا صفيحة، ولكنها أطول، وتقع إما أفقيًا (في الثعابين) أو عموديًا (في السحالي). وفيها، عندما تزداد وظيفة المخيخ، تشكل الصفيحة طية عرضية، تنحني بطريقة ظهرية مقوسة (في بعض الزواحف)؛ في التماسيح يتم تقسيمها عن طريق الأخاديد العرضية الأولية والثانوية - التلم الأمامي والخلفي - إلى فصوص أمامية ووسطى وخلفية. ترتبط الندفات المذكورة أعلاه بالأخيرة (الشكل 161-4).
في الطيور وخاصة في الثدييات، بالإضافة إلى تلك المشار إليها، تظهر أخاديد عرضية جديدة مع الحفاظ على النوع الرئيسي لهيكل الجسم المخيخي على شكل صفيحة مقوسة في الاتجاه الظهري. بفضل الأخاديد، فإن سطح جسم المخيخ يتخلله جيري عرضية، والتي لها أسماء خاصة في الثدييات. يُظهر القسم السهمي من المخيخ النخاع الأبيض، الذي يخترق كل تلفيف على شكل فروع شجرة؛ يُطلق على النمط العام للمادة البيضاء اسم شجرة الحياة - سيرة الشجرة (الشكل 183-5).
في الثدييات، في الفص الأوسط، بالإضافة إلى الأخاديد المستعرضة، هناك أخاديد طولية أخرى، تفصل بين الجزء الأوسط غير المتزوج - الدودية - والفصوص الجانبية، أو نصفي الكرة المخيخية - Bemisphaerae cerebelli (الشكل 181-7، 8؛ 161-أ، 5). لا يزال نصفي الكرة المخيخية في أحاديات المسلك واضحًا قليلاً؛ يصلون إلى أقصى تطور لهم في الثدييات الأعلى بسبب ظهور القدرة على الحركات المنعزلة للأطراف وظهور الروابط بين المخيخ والقشرة الدماغية. لذلك، في الثدييات، يبدأ الجزء البطني من الدماغ المؤخر، البونس (فارولي) (الشكل 175-15)، في البروز، مما يمثل تقاطعًا في مسارات التوصيل، وهو أمر مهم بشكل خاص عند البشر، الذين لديهم أقوى قشرة دماغية و أعلى قدرة على حركات الأطراف المعزولة. يقع الجسر على شكل سلسلة من التلال المستعرضة أمام النخاع المستطيل. بنهاياته الجانبية يغوص في المخيخ ليشكل أرجله الجانبية أو الجانبية - العضدية المخيخية الوحشية.

بسبب تجانس وظيفة المخيخ، فإن بنيته النسيجية موحدة (الشكل 162). بالفعل في حالة بدائية (في جلكيات)، يتكون النخاع الرمادي، الذي يشكل قشرة المخيخ، من ثلاث طبقات رئيسية من الخلايا: الخلايا السطحية الجزيئية، والخلايا العميقة الحبيبية، والخلايا المتوسطة الكبيرة. الأخيرة في سلسلة الحيوانات شديدة التمايز وتعرف باسم خلايا بوركينجي ذات التشعبات الشبيهة بالشجرة في الطبقة الجزيئية السطحية بدقة في المستوى السهمي للجسم، بينما تتفرع التشعبات من خلايا الطبقة الحبيبية في الطبقة القطاعية طائرة. بالإضافة إلى القشرة، يوجد في مخيخ الثدييات أيضًا مجموعات خاصة من الخلايا التي تشكل نواة المخيخ، وأهمها النواة المسننة المقترنة - النواة المسننة - الموجودة في نصفي الكرة المخيخية (الشكل 163-18). ). يوجد أيضًا عدد من النوى المزدوجة الموجودة بين النوى المتعرجة. تتلقى القشرة المخيخية نبضات من الحبل الشوكي والنخاع المستطيل، ومن الزيتون، والنوى السمعية للجسر والقشرة الدماغية. يتم إرسال الخلايا العصبية للخلايا القشرية إلى نوى المخيخ (المسنن وغيرها) ومن هنا إلى الدماغ المتوسط (النواة الحمراء)، إلى الجسر وإلى النخاع المستطيل. من خلال هذه المراكز، يتم إرسال النبضات من المخيخ عبر الحبل الشوكي إلى الأطراف.

تشكل جميع المسارات ثلاثة أزواج من السويقات المخيخية: من الجمجمة إلى الدماغ المتوسط، ومن الجانب إلى الجسر والذيلية إلى النخاع المستطيل، وهو ما يتوافق مع تقسيم المخيخ إلى ثلاثة فصوص رئيسية - الأمامي والوسطى والخلفي.
الدماغ المتوسط
الدماغ المتوسط - الدماغ المتوسط (الشكل 159 و160-3) - في الحيوانات السفلية، وفي الحالة الجنينية في الحيوانات العليا يصل إلى أحجام كبيرة مقارنة بأجزاء أخرى من الدماغ. ومع ذلك، في أشكال البالغين الأعلى يكون أقل تطورًا بكثير. فقط في الجلكيات، يتم تمثيل قبو الدماغ المتوسط جزئيًا بواسطة صفيحة ظهارية، وفي أسماك الجريث وجميع الفقاريات الأخرى، يتكون القبو - السقف البصري - من الأنسجة العصبية التي تنتجها الصفيحة الجانبية الجنينية. نتيجة لذلك، يتم تقليل تجويف الدماغ بشكل كبير، ويتحول في أشكال أعلى إلى قناة قناة الدماغ التي تربط البطين الدماغي الرابع من الدماغ المعين مع البطين الدماغي الثالث من الدماغ البيني. الحدود الأمامية للدماغ المتوسط عبارة عن حزمة من الألياف العصبية - الصوار الخلفي - الصوار الخلفي. في الحيوانات السفلية، يوجد في القبو حديبتان أو فصان بصريان، lobi Opticali، وهما مركز الرؤية؛ تنتهي فيها ألياف العصب البصري. في الزواحف، جزئيًا، وفي الثدييات، تنتهي جميع ألياف العصب البصري تقريبًا في الدماغ البيني، وبالتالي يتم تقليل الفصوص البصرية مقارنةً بالمهاد البصري (الشكل 164). في الوقت نفسه، حتى في البرمائيات، ترتبط الفصوص الخلفية تشريحيا بجهاز السمع والتوازن. يتم تمييز هذه الفصوص بشكل عياني بشكل أكثر وضوحًا في بعض الزواحف والطيور وخاصة في الثدييات التي لا تحتوي على أكيمة، ولكن رباعية التوائم - الأجسام الرباعية (الشكل 182-20، 21). في بعض الثدييات التي تتمتع بسمع جيد، تكون الأكيمة الخلفية أكبر من الأكيمة الأمامية (في الحيتان والحيوانات آكلة اللحوم) (الشكل 188-6)، ولكن عادة ما تكون الفصوص البصرية الأمامية أكبر من الفصوص الخلفية.
بسبب المادة الرمادية للوحة الرئيسية الجنينية، الموجودة في الجزء البطني من تجويف الدماغ المتوسط، يتم تشكيل الغطاء - سقيفة السويقات (الشكل 187-3)، ويتكون من نوى المحرك للعين والأعصاب البُكرية - نوى العصب الثالث والرابع - والنواة الحركية للغطاء هي النواة الحركية. هذا الأخير بمثابة استمرار لكتلة خلوية مماثلة لمركز النخاع المستطيل، كما يمكن رؤيته في دماغ الحيوانات الأكثر بدائية. بدءًا من الزواحف، فهو يتكون من عدد من النوى الفردية. وأهمها النواة الحمراء، النواة روبر (7)؛ فهو ينهي الألياف العصبية التي تشكل السويقة الأمامية للمخيخ. أما النواة الأخرى بين السويقتين - النواة بين السويقتين - فهي متصلة بالعضو الشمي من خلال عقدة اللجام.
من الناحية البطنية بين المخ والحبل المعيني والحبل الشوكي، عند قاعدة الدماغ المتوسط في الثدييات، توجد طبقة من المسارات - سيقان المخ - ريدونكولي المخي (الشكل 175-16. 187-5). هم أكثر قوة في الثدييات العليا. والذي يرتبط بتطور العباءة. في الأجزاء الوسطى من الساقين توجد مسارات حركية، وفي الأجزاء الجانبية توجد مسارات حسية.

يتلقى الدماغ المتوسط نبضات من المخيخ وأجهزة الرؤية، وفي الحيوانات الأرضية، بالإضافة إلى ذلك، من كارا نصفي الكرة المخية ومن أجهزة السمع والتوازن، فإنه يرسل نبضات إلى الحبل الشوكي وإلى الحبل المتوسط.
الدماغ البيني
يحتل الدماغ البيني (الشكل 159 و160-2) مساحة كبيرة إلى حد ما من الدماغ مع تجويف كبير في البطين الثالث. ولكن بعد ذلك، يصبح التجويف البطيني شبيهًا بالشق.
تعمل الصفيحة السقيفية بمثابة قبو للبطين الثالث، والذي يظل بدائيًا في جميع الحيوانات، ويتكون من صفيحة ظهارية - الصفيحة الظهارية - والتي، مندمجة (كما هو الحال في الدماغ المعيني) مع الأم الحنون، تشكل السقف الوعائي للبطين الثالث البطين الدماغي - tela chorioidea ventriculi tertii، الذي يحتوي على الضفيرة المشيمية. يتم إدخال السقيفة عن طريق العمليات في تجويف البطين الثالث، ومن خلال الثقبة بين البطينات تخترق أيضًا الدماغ الانتهائي، حيث تمر إلى الضفائر المشيمية للبطينات الدماغية الجانبية (الشكل 186-4، 10)، والتي تشكلت بسبب الصفيحة القطبية للدماغ الانتهائي.
مشتقات القبو هي: 1) نمو غير مزدوج على شكل أنبوب - المشاش و 2) عقدة مقترنة من اللجام.
المشاش، أو الغدة الصنوبرية، - المشاش (الشكل 159-6) هو بداية للثالثة، ما يسمى بالعين الجدارية. الغدة الصنوبرية، الموجودة في جميع الحيوانات تقريبًا، ليست متطورة بشكل متساوٍ في جميع الحيوانات وتغيب فقط في عدد قليل من الحيوانات (الجرابيات وبعض الحيوانات الأخرى).
في الفقاريات السفلية، لا يلعب الدماغ البيني هذا الدور كما هو الحال في السلى، وهذا هو السبب في أنه متطور نسبيًا فيها. فقط مع حركة المراكز البصرية من الدماغ المتوسط \u200b\u200bإلى داخلها (الشكل 164)، بسبب الانتقال إلى نمط الحياة الأرضي، يبدأ الدماغ البيني في الزيادة، تاركا الدماغ المتوسط \u200b\u200bبعيدا، وهو أمر ملحوظ بشكل خاص في البشر. نظرًا لوجود عدد كبير من نوى المادة الرمادية، يصبح الدماغ البيني مركزًا للارتباط للعديد من المسارات المؤدية إلى القشرة الدماغية والعودة؛ ومن هنا فمن الواضح أن تمايز الدماغ البيني يبدأ من لحظة نمو الدماغ الانتهائي.
يحتوي الدماغ البيني على عدد من المراكز النباتية: الحرارية، الحركية الوعائية، الإفرازية، استقلاب البروتين والكربوهيدرات، الماء، إلخ.
الدماغ المحدود
الدماغ الانتهائي (الشكل 159 و160-1، 9) هو جزء من الدماغ يتطور بقوة شديدة في عدد من الحيوانات ويصل إلى أعلى مستويات تطوره في فئة الثدييات، وخاصة في الرئيسيات. في البشر، يتفوق الدماغ الانتهائي على جميع أجزاء الدماغ الأخرى لدرجة أنه يغطيها كلها تقريبًا. ينقسم الدماغ الانتهائي إلى الدماغ الشمي - الدماغ الأنفي - والباليوم - الباليوم.
الدماغ الشمي
من الجدار الأمامي للدماغ الانتهائي ينمو زوج من النتوءات - البصيلات الشمية - البصلة الشمية (الشكل 159-10)، - تتصل بالدماغ بقاعدة عريضة أو أرجل طويلة أكثر أو أقل - المسالك الشمية - السبيل الشمي (في العظام الأسماك والزواحف).
تأخذ القاعدة، أو الجزء القاعدي، من الدماغ أبعادًا مهمة موجودة بالفعل في الدماغ البدائي. إنه بمثابة مركز شم ثانوي ويتكون من سماكة كبيرة - العقدة القاعدية - النواة القاعدية (1) ، والتي يبرز منها الجسم المخطط - الجسم المخطط - في تجويف البطينين الجانبيين للدماغ. ينقسم النخاع الرمادي للمخطط أثناء التطور بواسطة النخاع الأبيض، الذي يشكل المحفظة الداخلية - المحفظة الداخلية (الشكل 184-9)، - إلى الجزء الظهري، أو الجسم المذنب - النواة المذنبة، - والجزء البطني، أو نواة عدسية الشكل (2) -nucleus lentiformis، والمحفظة الخارجية -capsula externa (4) - تفصل الجزء الجانبي، أو السياج (3)، -claustrum. بسبب النمو القوي، تضغط الأجسام المهاجمة على تجويف البطينين الجانبيين، وتحولها إلى مساحات تشبه الشق (5).
في دماغ الثدييات شديد التمايز، يتطور الجزء الشمي منه في عدد من الحيوانات البعيدة عن التماثل.
في الحيوانات الكبيرة الحجم (وهذا يشمل جميع الحيوانات الأليفة، وبشكل عام، معظم الحيوانات التي لديها حاسة شم متطورة)، يكون الجزء الشمي من الدماغ مهمًا جدًا.
في الحيوانات المجهرية (القرود والحيتان المسننة والدلافين وحيتان العنبر) يكون الجزء الشمي من الدماغ صغيرًا وفقًا لعمل العضو الشمي.
من بين الأجزاء الفردية من الدماغ الشمي، تكون المصابيح الشمية بارزة بشكل خاص، والتي تكون في الحيوانات الكبيرة الحجم زوائد للدماغ بارزة من الأمام (الشكل 177-I)، وفي الحيوانات المجهرية تكون صغيرة جدًا. يتشكل الجزء الخارجي (البطني) من العقدة القاعدية، أو القشرة الشمية، أمام التصالب البصري، بين المسالك الشمية الجانبية والوسطى، الحديبة الشمية، والتي تكون أكثر وضوحًا بين الحيوانات الأليفة فقط في الخنازير؛ وفي حالات أخرى يكون مسطحًا، ونتيجة لذلك يطلق عليه ببساطة المثلث الشمي. في الحيوانات المجهرية، المنطقة المقابلة بالكاد تبرز.
هيكل القشرة الدماغية
العباءة الثانوية مصنوعة من مادة الدماغ الرمادية والبيضاء. الأول يقع بشكل سطحي ويشكل القشرة الدماغية، والثاني يقع في العمق.
إن القشرة الدماغية في أجزائها المختلفة ذات بنية مختلفة من الناحية النسيجية، وهو الأمر الذي أسسه لأول مرة V. A. Betz (1874) وأوضح أيضًا: “إن هذا التمايز الهيكلي للقشرة الدماغية هو تعبير عن توطين الوظائف في القشرة” (1881) . يتعلق الاختلاف بموقع العناصر الخلوية (البنية الخلوية) ومسار الألياف في النخاع الرمادي (البنية النخاعية). تمت دراسة البنية الخلوية والنخاعية للقشرة الدماغية بتفاصيل أكثر أو أقل فقط عند البشر وبشكل سيء إلى حد ما عند الحيوانات.
تقع العناصر الخلوية للقشرة بالتوازي مع سطح الدماغ في ست طبقات (طبقات). هذه الطبقات، التي تحسب من الخارج إلى الداخل، هي كما يلي: الطبقة الأولى الجزيئية، الطبقة الثانية الحبيبية الخارجية، الطبقة الثالثة من الخلايا الهرمية الصغيرة، الطبقة الرابعة الحبيبية الداخلية، الطبقة V من الخلايا الهرمية الكبيرة والطبقة السادسة من الخلايا متعددة الأشكال ( الشكل 166). في سلسلة النشوء والتطور للحيوانات، لم يتم تحديد جميع الطبقات الست بشكل حاد، ولكن يمكن إثبات وجودها في جميع الثدييات.

تنقسم الخلايا التي تشكل القشرة إلى ثلاثة أنواع: رامون-كاخاليفسكي، والهرمية، ومتعددة الأشكال. من بين هذه الخلايا الهرمية في بيتز، تستحق اهتماما خاصا: كلما كان الحيوان أعلى في التنظيم أو أكبر سنا، كلما زادت العمليات التي لديهم، وبالتالي، فإن اتصالات هذه الخلايا مع الآخرين تكون أكثر اتساعا (الشكل 167).
يتم تعيين وظائف مختلفة لطبقات الخلايا المختلفة: الطبقة الحبيبية الداخلية (IV)، وهي الطبقة الأساسية، لها وظيفة مستقبلات، وخلايا الطبقتين V و VI، والخلايا الهرمية الكبيرة والمغزلية لها وظيفة المستجيب. لاحقًا، تؤدي الخلايا الظاهرة في الطبقتين II وIII وظائف ارتباط ذات مستوى أعلى، أي وظائف ذهنية.
على أساس الاختلافات المحلية في الهندسة المعمارية الخلوية، تنقسم قشرة الثدييات إلى مناطق رئيسية - مجالات - مناطق - وكل مجال، على الأقل في الثدييات العليا وخاصة في البشر، على أساس الهندسة المعمارية النخاعية المختلفة يمكن تقسيمها إلى عدد من المناطق المعمارية النخاعية.
من الناحية التطورية، يتم تمييز جميع الحقول الموجودة في العباءة الثانوية عن المناطق الأربع البدائية للثدييات السفلية (الجرابيات والحيوانات آكلة الحشرات)؛ تختلف هذه المناطق في هندستها الخلوية، واستنادا إلى وجود بعض ألياف الإسقاط فيها (الشكل 165)، فإنها تتوافق أيضا مع مراكز وظيفية خاصة. وهكذا، في المنطقة الأمامية يقع المركز القشري الحركي البدائي، وفي الخماس البصري القذالي، بينهما الخماس الحسي الجلدي، وفي المنطقة الزمنية هو الخماس السمعي. لا تتوافق المراكز الفسيولوجية والمعمارية دائمًا مع موقع الجيري.
يتم تحديد نسبة ألياف اللب في القشرة الدماغية إلى حد ما من خلال التقسيم المقبول لها إلى مجموعات تتوافق مع ست طبقات من الخلايا (الشكل 168، على اليمين).

بناءً على تطور غمد المايلين (النقوي الوراثي)، يتم تصنيف جميع الألياف العصبية إما كنظام إسقاط أو نظام ارتباط. الأول يتطور قبل الولادة، والثاني بعد الولادة فقط. من الناحية الوراثية، يمكن أيضًا تقسيم القشرة وفقًا لخاصيتين - النسبة المئوية للألياف البارزة والارتباطية وتكوينها - إلى مناطق: أولية ونهائية ومتوسطة، أي الألياف المغطاة بغمد المايلين قبل الولادة وبعد الولادة وفي الفترة الأخيرة من الحالة الجنينية.
حاليًا، استنادًا إلى دراسة البنية الخلوية والنخاعية للقشرة الدماغية، تم تحديد أكثر من 250 مجالًا لدى البشر.
تربط الألياف الرابطة مناطق عديدة من القشرة مع بعضها البعض داخل نصف الكرة المخية الواحد. وهي مقسمة إلى قصيرة وطويلة. الأول يربط التلافيف المجاورة، والأخير يربط المناطق والفصوص الأكثر بعدًا عن بعضها البعض. يتم تطوير الألياف الطويلة بشكل خاص عند البشر، بينما تكون في الحيوانات إما ضئيلة أو غائبة تمامًا.
تربط الألياف الصوارية المناطق الموجودة في نصفي الكرة الأرضية المختلفين. إنهم يشكلون الصوار الأمامي والخلفي (الصوار) - صوار قرن عمون والجسم الثفني.
تنقسم الألياف الإسقاطية تشريحيًا إلى قصيرة وطويلة، ووظيفيًا إلى قشرية قشرية وبتلة قشرية. تربط ألياف الإسقاط القصيرة القشرة بنواة الأكيمة البصرية والقناة الرباعية التوائم. تربط المسالك الطويلة القشرة بالنواة الجسرية أو بالحبل الشوكي. كل هذه المسارات، القصيرة والطويلة، تمر بين الجسم المذنب ونواة التلال البصرية من ناحية، والنواة العدسية من ناحية أخرى، وتشكل المحفظة الداخلية - المحفظة الداخلية (الشكل 184-9). . في الدماغ المتوسط، تشكل هذه الألياف السويقة الدماغية.
مسارات الإسقاط، وكذلك مسارات الارتباط، هي الأكثر تطورًا عند البشر؛ في الحيوانات، بعضها غائب تماما.
وبالتالي، فإن نصفي الكرة المخية بشكل عام والقشرة الدماغية بشكل خاص يتم تنظيمهما بشكل أكثر تعقيدًا، علاوة على ذلك، بشكل غير متساوٍ بين الثدييات، حيث يصلان إلى أقصى تطور في الرئيسيات العليا وخاصة عند البشر. وفقا ل I. P. Pavlov، فإن نصفي الكرة المخية تتعرض باستمرار لقصف لا تعد ولا تحصى من العالم الخارجي ومن البيئة الداخلية للجسم نفسه. وبما أن القشرة الدماغية للحيوانات العليا هي الحاملة لوظيفة الإغلاق (أي وظيفة الاكتساب، وتكوين روابط جديدة بين الكائن الحي والبيئة، وتطوير تجارب حياتية جديدة، ووظيفة التكيف الجيني الذي يتكيف مع الكائن الحي) للظروف البيئية)، ويتولى القسم الأعلى للجهاز العصبي المركزي معرفته بجميع الظواهر التي تحدث في الجسم.
وزن الدماغ
يختلف وزن الدماغ المطلق للحيوانات بشكل كبير. ويتم تحديده حسب حجم الحيوان وعمره وفصيلته وما إلى ذلك. وتوجد أكبر الأدمغة في الحيوانات الكبيرة. يصل وزنها إلى: للحيتان - 4673-7000.0، للفيلة - 4370-5430.0، للخيول - 372-570.0، للماشية - 410-550.0، في الأغنام - 97 -112.0، في الخنازير - 96-145.0، في الكلاب - 46 -138.0 في الفئران - 0.37. الوزن المطلق للدماغ عند البشر مرتفع جدًا - 1350-1450.0.
يتناسب الوزن النسبي للدماغ بشكل عام عكسيًا مع وزن الحيوان: في الحيتان فقط 1: 10590 وحتى 1: 14000، في الفيلة - 1: 375-560، في الخيول - 1: 480-1000 (1)، في الخنازير - 1: 1200-1900، في الكلاب - 1: 30-400. في البشر الوزن النسبي للدماغ هو 1: 35-45. في الحيوانات الصغيرة، يكون الوزن النسبي للدماغ أعلى بكثير منه عند البالغين؛ في الحيوانات الصغيرة يكون وزن الدماغ كبيرًا جدًا: في القرد العنيد المظلم يصل إلى 1:15.
نسبة وزن الحبل الشوكي إلى الدماغ: عند البشر - 1: 40، في الخيول - 1: 2.5، في الثيران - 1: 1.5، في الكلاب - 1: 4.5-9. وبذلك يبلغ وزن الحبل الشوكي عند الإنسان 2.5% فقط من وزن الدماغ، وفي الخيول 40%.
المادة الرمادية أكثر في الحيوانات الصغيرة، فمثلا نسبة المادة الرمادية إلى المادة البيضاء هي 52.1 و 47.9 في الخيول، 50.1 و 49.9 في الماشية، 61.1 و 38.9 في الكلاب، وحتى النمس لديه 80.7 و 19.3 (بوبوف). .






