Ciclo mestruale- cambiamenti ciclici nel corpo di una donna, la cui manifestazione esterna sono le mestruazioni.
I cambiamenti ciclici nelle ovaie - il ciclo ovarico - si dividono in fasi follicolari e luteiniche, mentre i cambiamenti nell'endometrio - il ciclo uterino - in fasi proliferative e secretorie. Come risultato del rigetto dello strato funzionale dell'endometrio, si verificano le mestruazioni. Il menarca è la prima mestruazione. Di solito si osserva a 10-12 anni di età, mentre un ciclo regolare si instaura solitamente dopo 1-1,5 anni. Il ciclo medio è di 28 giorni, quello normale va da 21 a 35 giorni. Il primo giorno delle mestruazioni corrisponde al primo giorno del ciclo mestruale. La durata delle mestruazioni è di 2-7 giorni (in media 4-5 giorni), la perdita di sangue va da 50 a 150 ml (in media 70-100 ml).
Il ciclo mestruale è determinato dal lavoro coniugato di cinque anelli della catena neuroumorale (corteccia cerebrale, ipotalamo, ghiandola pituitaria, ovaie, utero).
I principali prodotti di secrezione dell'ipotalamo sono i fattori di rilascio ipofisari. L'ormone di rilascio delle gonadotropine (GnRH) controlla la secrezione delle gonadotropine ipofisarie, degli ormoni luteinizzanti (LH) e degli ormoni follicolo-stimolanti (FSH) (Fig. 4).
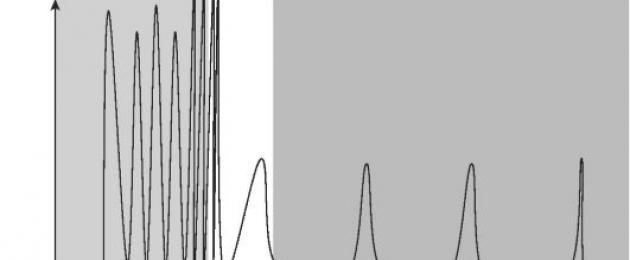
Il GnRH è l'unico ormone che regola la secrezione pulsante di due ormoni ipofisari. L'infusione cronica di GnRH non stimola la secrezione di gonadotropine. La modalità pulsatile della secrezione di GnRH (Fig. 5) è necessaria a causa del periodo molto breve di puluralisi RG - circa 2-4 minuti. Durante il ciclo mestruale, la frequenza e l'ampiezza delle pulsazioni del GnRH cambiano: nella fase follicolare sono elevate e nella fase luteinica diminuiscono.
Riso. 4. Regolazione del ciclo mestruale
 Riso. 5. Modello di secrezione del GnRH
Riso. 5. Modello di secrezione del GnRH
La fine di ogni ciclo mestruale e l'inizio di quello successivo sono caratterizzati da bassi livelli di steroidi sessuali: progesterone ed estrogeni.
Con la cessazione della funzione del corpo luteo aumenta la produzione di FSH e LH. Le cellule della granulosa interagiscono con l'FSH e le cellule dello strato tecale interno interagiscono con l'LH. Ogni ciclo mestruale(Fig. 6) da 3 a 30 follicoli primordiali, sotto l'influenza dell'FSH, entrano nella fase di crescita e secernono estrogeni, il cui livello aumenta progressivamente durante la 1a - fase follicolare ciclo mestruale.
Durante la crescita dei follicoli secondari (entro l'8° giorno del ciclo mestruale), dominante follicolo che si trasforma in follicolo terziario(preovulatoria, di seguito grafici bolla, fino a 2-3 cm di diametro).
La sintesi degli estrogeni si ottiene in due modi. Il primo percorso prevede l'aromatizzazione enzimatica degli androgeni in estrogeni da parte delle cellule della granulosa. La seconda via è associata alla sintesi degli estrogeni nelle cellule tecali nelle ultime fasi della fase antrale. Pertanto, a metà della fase follicolare, aumenta il livello di estrogeni e androgeni follicolari, accompagnato da una diminuzione della concentrazione di FSH (feedback negativo).
Allo stesso tempo, gli estrogeni stimolano la secrezione di LH durante tutto il periodo follicolare.
L'FSH stimola:
Fase di crescita dei follicoli primordiali;
Trasporto di liquido nella cavità follicolare;
Espressione dei recettori per LH e acido prolattico sulle cellule della granulosa;
Attività dell'aromatasi. L'LH stimola:
Produzione da parte delle cellule follicolari di proteine a basso peso molecolare che neutralizzano il fattore che sopprime la meiosi;
Divisione meiotica dell'ovocita e transizione allo stadio del 2o ordine - set aploide;
Sintesi degli androgeni - androstenedione e testosterone - nelle cellule il CA;
Sintesi del progesterone (luteinizzazione) nelle cellule follicolari;
Sintesi delle prostaglandine nelle cellule follicolari;
Induzione dell'ovulazione.
Nella fase preovulatoria, sulle cellule della granulosa del follicolo, l'FSH induce lo sviluppo di recettori per LH e prolattina. Pertanto, alla fine della fase preovulatoria, il contenuto di FSH e LH aumenta e le cellule follicolari diventano insensibili agli effetti degli estrogeni e degli androgeni. Alte concentrazioni di estrogeni stimolano il rilascio di LH e la rottura della parete della vescicola di Graaf (follicolo terziario) - cioè ovulazione, che si verifica 10-12 ore dopo il picco dei livelli di LH. Quindi l'uovo viene rilasciato nella cavità addominale e inizia fase luteinica del ciclo.
Estrogeni:
Stimolare la proliferazione delle cellule follicolari;
Stimolare l'espressione dei recettori dell'FSH;
Partecipano (insieme all'FSH) alla formazione dei recettori dell'LH nelle cellule follicolari;
Aumentare la secrezione di LH; con un alto contenuto di estrogeni, il GnRH stimola le cellule che sintetizzano LH;
Sopprime la secrezione di FSH; quando i livelli di estrogeni sono bassi, il GnRH stimola le cellule che sintetizzano l’FSH.
Androgeni:
Inibire l'espressione dei recettori dell'FSH sulle cellule della granulosa;
Inibisce l'attività dell'aromatasi.
I capillari risultanti crescono rapidamente nella cavità del follicolo, le cellule della granulosa subiscono la luteinizzazione sotto l'influenza dell'LH, che porta alla formazione del corpo luteo.
I livelli di estrogeni iniziano a diminuire a partire dalla fine della fase preovulatoria in un contesto di elevate concentrazioni di FSH e LH, continuano a diminuire durante la fase luteinica iniziale e aumentano nuovamente a causa della secrezione del corpo luteo.
corpo luteo(corpo luteo)è una ghiandola endocrina transitoria che funziona per 8-14 giorni, indipendentemente dalla durata del ciclo mestruale, e sintetizza progesterone, estrogeni (principalmente 17b-estradiolo) e prolattina. I livelli di progesterone aumentano gradualmente dopo l’ovulazione e raggiungono il picco 8-9 giorni dopo l’ovulazione, che corrisponde approssimativamente al momento dell’impianto. L'effetto termogenico del progesterone porta ad un aumento della temperatura corporea di almeno 0,33°C (l'effetto dura fino alla fine della fase luteinica).
Progesterone:
Prepara l'endometrio all'annidamento;
Rilassa le fibre miometriali;
Ha un effetto natriuretico, stimolando la secrezione di aldosterone;
Il progesterone placentare viene metabolizzato nella corteccia surrenale fetale e nei testicoli come precursore rispettivamente dei corticosteroidi e del testosterone.
Pertanto, la fase luteinica è caratterizzata da un aumento delle concentrazioni di progesterone e prolattina e da bassi livelli di FSH e LH.
Quando la funzione del corpo luteo regredisce, le concentrazioni di steroidi sessuali diminuiscono e inizia un nuovo ciclo mestruale.
Oltre agli ormoni elencati, il corpo luteo e successivamente la placenta producono relaxina. Inibisce l'attività contrattile del miometrio attivando l'azione del progesterone e aumentando il livello di cAMP sia nelle cellule muscolari lisce del miometrio che nei condrociti della sinfisi pubica, provocandone l'ammorbidimento.
Corpo bianco- cicatrice del tessuto connettivo al posto della funzione completata e corpo luteo degenerato.
CICLO OVARICO
Il numero di oogoni in un embrione femminile a metà dello sviluppo intrauterino raggiunge i 5-7 milioni, tuttavia, una parte significativa degli ovociti subisce atresia (Fig. 7), associata a una bassa produzione di ormoni gonadotropici. Le ovaie di una neonata contengono già 1-2 milioni di ovociti; entro il periodo della pubertà ce ne sono da 100 a 400mila.Durante il periodo riproduttivo, il 98% dei follicoli primordiali muore, circa il 2% raggiunge lo stadio di follicoli primari e secondari, ma non più di 400-500 ovulano. Tutti i follicoli che hanno iniziato a svilupparsi ma non hanno raggiunto la fase di ovulazione vanno incontro ad atresia.
Pertanto, la durata della vita di un ovocita può raggiungere i 40-50 anni. Questo è il motivo per cui il rischio di difetti genetici fetali aumenta significativamente con l’età materna.
 Riso. 7. Modello di secrezione del GnRH
Riso. 7. Modello di secrezione del GnRH
STRUTTURA FOLLICOLARE
Follicolo primordiale ricoperto da un unico strato di cellule follicolari (granulosa) e circondato da una membrana basale. Le cellule della granulosa che circondano l'ovocita (“corona radiata”) secernono un substrato glicoproteico, che forma una zona trasparente - zona pellucida- tra l'ovocita e le cellule della granulosa. Sulla superficie della zona pellucida sono presenti recettori specie-specifici per l'interazione solo con gli spermatozoi allogenici; la penetrazione della zona da parte di uno spermatozoo porta allo sviluppo di una “reazione zonale” che impedisce la polispermia.
Le cellule dello stroma ovarico formano uno strato di cellule fusate - teca. Gli androgeni, che assicurano lo sviluppo dei follicoli, sono prodotti esclusivamente dalle cellule tecali. Come risultato della proliferazione, questi ultimi si dividono in due strati: quello interno, che ha una struttura ghiandolare, e quello esterno - tessuto connettivo. Tra di loro si accumula il liquido follicolare, contenente il trasudato sierico e la secrezione mucopolisaccaridica delle cellule della granulosa. L'accumulo di liquido conferisce al follicolo l'aspetto di una vescicola e viene chiamato tale follicolo
è secondario o antrale (Fig. 8). L'ovocita in esso contenuto non ha ancora subito la seconda divisione meiotica.
Il follicolo primordiale è ricoperto da un unico strato di cellule follicolari (granulosa) e circondato da una membrana basale
 Riso. 8. Crescita del follicolo
Riso. 8. Crescita del follicolo
 Riso. 9. Follicolo terziario (“vescicola di Graaf”)
Riso. 9. Follicolo terziario (“vescicola di Graaf”)
Lo sviluppo dell'ovocita continua fino alla fecondazione e la trasformazione di un ovocita del 1° ordine in un ovocita del 2° ordine, che ha già un corredo cromosomico aploide, avviene immediatamente prima dell'ovulazione o nelle tube di Falloppio.
Durante ogni ciclo ovarico, nelle ovaie si sviluppano 15-20 follicoli. Alcuni di essi raggiungono grandi dimensioni (fino a 8 mm) entro la metà del ciclo, ma solo un follicolo diventa un follicolo terziario maturo con un diametro di 2-3 cm (“vescicola di Graaf”, Fig. 9).
CICLO UTERINO
I cambiamenti nei livelli ormonali influenzano direttamente le condizioni dell'endometrio, della mucosa delle tube di Falloppio, del canale cervicale e della vagina. La mucosa uterina subisce cambiamenti ciclici (fasi proliferativa, secretoria e mestruale). L'endometrio è diviso in strati funzionali (che scompaiono durante le mestruazioni) e basali (che si conservano durante le mestruazioni).
Fase proliferativa- la prima metà del ciclo - dura dal primo giorno delle mestruazioni fino al momento dell'ovulazione. È caratterizzato dalla rigenerazione dello strato funzionale dovuta alla migrazione e alla proliferazione delle cellule epiteliali delle ghiandole dello strato basale verso la superficie sotto l'influenza degli estrogeni (principalmente estradiolo). Nell’endometrio si formano nuove ghiandole uterine e dallo strato basale crescono le arterie spirali. La durata della fase può variare. La temperatura corporea basale è normale.
Fase secretoria- seconda metà - dura dall'ovulazione all'inizio delle mestruazioni. Le cellule epiteliali smettono di dividersi e di ipertrofizzarsi. Le ghiandole uterine si espandono e le cellule ghiandolari secernono attivamente glicogeno, glicoproteine, lipidi e mucina. Nelle parti superficiali dello strato funzionale aumenta il numero delle cellule del tessuto connettivo attorno alle quali si formano fibre collagene e reticolari. Arterie a spirale diventano più contorte, avvicinandosi alla superficie della mucosa. Se l'impianto dell'ovulo fecondato non si è verificato, una diminuzione del contenuto degli ormoni steroidei ovarici porta alla torsione, alla sclerosi e alla diminuzione del lume delle arterie spirali che forniscono i due terzi superiori dello strato funzionale dell'endometrio. Di conseguenza, si verifica un deterioramento del flusso sanguigno nello strato funzionale, ischemia e rigetto, ad es. sanguinamento.
fase mestruale- rigetto dello strato funzionale dell'endometrio, dura 3-7 giorni.
Per determinare il momento dell'ovulazione vengono utilizzati vari metodi di diagnostica funzionale delle fasi del ciclo mestruale. Sono definiti i seguenti parametri.
1. Temperatura basale. È associato all'effetto termogenico del progesterone (Fig. 10).
Giorno del ciclo 1 23456789 1010 11 12 13 14 15 16 17 1819 20 22 23 24 25 2627 2829
 Riso. 10. Temperatura basale (rettale).
Riso. 10. Temperatura basale (rettale).
2. Estensibilità del muco cervicale. Sotto l'influenza degli estrogeni, l'estensibilità del muco aumenta significativamente. Raggiunge i valori massimi durante l'ovulazione (Fig. 11)
3. L'effetto di arborizzazione del muco cervicale (fenomeno della “felce”). Questo fenomeno è più pronunciato durante il periodo di ovulazione a causa di
un'elevata concentrazione di sali di sodio che precipitano in cristalli (sintomo di cristallizzazione), somiglianti esternamente ad una superficie a forma di albero o felce (Fig. 12).
4. Indice cariopinotico - KPI (utilizzando l'analisi microscopica di uno striscio vaginale).
 Riso. undici. Il grado di estensibilità del muco cervicale
Riso. undici. Il grado di estensibilità del muco cervicale
 Riso. 12. Il fenomeno della "felce".
Riso. 12. Il fenomeno della "felce".
Il CPI è il rapporto tra le cellule cheratinizzate con nuclei picnotici (punteggiati) e tutte le cellule dell'epitelio vaginale nello striscio (Fig. 13, vedere riquadro). Il valore CPI più alto corrisponde al periodo di ovulazione - 70-80%, nei restanti giorni del ciclo mestruale - fino al 30-40%.
Embriologia- la scienza dell'embrione, le leggi del suo sviluppo. L'embriologia medica studia i modelli di sviluppo dell'embrione umano, le caratteristiche strutturali, metaboliche e funzionali della barriera placentare (sistema madre-placenta-feto), le cause di deformità e altre deviazioni dalla norma, nonché i meccanismi di regolazione dell'embriogenesi.
Il concetto di embriogenesi comprende il periodo che va dal momento della fecondazione alla nascita (per gli animali vivipari), la schiusa delle uova (per gli animali ovipari) e la fine della metamorfosi (per gli animali con lo stadio di sviluppo larvale).
FECONDAZIONE
Trasporto dei gameti. Nell'uomo il volume normale dell'eiaculato è di circa 3 ml; contiene una media di 350 milioni di spermatozoi. Per garantire la fecondazione, il numero totale di spermatozoi deve essere almeno 150 milioni e la loro concentrazione in 1 ml deve essere almeno 60 milioni.Grazie all'elevata mobilità, gli spermatozoi in condizioni ottimali possono raggiungere la cavità uterina in 30 minuti - 1 ora, e in 1,5-2 ore si trova nella parte distale (ampollare) della tuba di Falloppio, dove avviene la fecondazione. Gli spermatozoi mantengono la capacità fecondante fino a 2 giorni.
L'ovocita di primo ordine rilasciato dall'ovaio durante l'ovulazione ha un diametro di circa 130 micron ed è circondato da una fitta zona pellucida, o membrana, e da una corona di cellule follicolari, il cui numero raggiunge i 3-4mila. risale dalle fimbrie della tuba di Falloppio (ovidotto) e si muove lungo di essa. Qui termina la maturazione della cellula germinale. In questo caso, a seguito della seconda divisione, si forma un ovocita (uovo) di secondo ordine, che perde i suoi centrioli e quindi la capacità di dividersi. Il nucleo di un ovulo umano contiene 23 cromosomi; uno di questi è il cromosoma sessuale X.
L'ovulo umano solitamente consuma la sua riserva di nutrienti entro 12-24 ore dopo l'ovulazione e poi muore se non viene fecondato.
Fecondazione si verifica nella parte ampollare dell'ovidotto. Condizioni ottimali per l'interazione dello sperma con l'uovo
solitamente creato entro 12 ore dopo l'ovulazione. Durante l'inseminazione, numerosi spermatozoi si avvicinano all'ovulo ed entrano in contatto con la sua membrana. L'uovo inizia a eseguire movimenti rotatori attorno al proprio asse ad una velocità di 4 rotazioni al minuto. Questi movimenti sono causati dal battito dei flagelli degli spermatozoi e durano circa 12 ore.Durante l'interazione delle cellule germinali maschili e femminili, in esse si verificano numerosi cambiamenti. Gli spermatozoi sono caratterizzati dai fenomeni di capacitazione e reazione acrosomiale. La capacitazione è il processo di attivazione degli spermatozoi nelle tube di Falloppio sotto l'influenza della secrezione mucosa delle cellule ghiandolari. Il progesterone attiva la secrezione delle cellule ghiandolari. Dopo la capacitazione segue una reazione acrosomiale durante la quale gli enzimi ialuronidasi e trypsin vengono rilasciati dagli spermatozoi. La ialuronidasi scompone l'acido ialuronico contenuto nella zona pellucida. La tripsina scompone le proteine del citolemma dell'uovo e delle cellule della corona radiata. Di conseguenza, le cellule della corona radiata si dissociano e la zona pellucida si dissolve.
Nell'uovo, il citolemma nell'area di attacco dello sperma forma un tubercolo di sollevamento, nel quale entra uno spermatozoo e appare una membrana densa - la membrana di fecondazione, che impedisce l'ingresso di altri spermatozoi e il fenomeno della polispermia. I nuclei delle cellule germinali femminili e maschili si trasformano in pronuclei, si avvicinano e inizia lo stadio synkaryon. Sorge zigote, ed entro la fine del 1° giorno dopo l'inizio della fecondazione frazionamento.
Il sesso del bambino dipende dai cromosomi sessuali del padre. A causa della maggiore sensibilità degli embrioni maschili agli effetti dannosi di vari fattori, il numero dei neonati maschi è inferiore a quello delle femmine: per ogni 100 maschi nascono 105 femmine.
Il movimento dell'uovo fecondato è assicurato dalle contrazioni peristaltiche dei muscoli del tubo e dallo sfarfallio delle ciglia dell'epitelio. L'embrione si nutre di piccole riserve di tuorlo nell'uovo e, eventualmente, del contenuto delle tube di Falloppio.
Il trasporto dell'embrione nell'utero avviene in un ambiente immunosoppressore, nella formazione del quale sperma, liquido di blastocisti, UN Proteina 2-uterina (inizia a essere prodotta dall'epitelio ghiandolare dell'endometrio nei giorni successivi all'ovulazione) e fattore precoce di gravidanza (EGF), descritto per la prima volta da H. Morton nel 1974. L'EGF viene prodotto dall'uovo 46-48 ore Dopo
le fecondazione ed è uno dei primi indicatori di gravidanza e il primo agente immunosoppressore che previene il rigetto della blastocisti. Fattori di protezione immunologica:
UN 2 - proteina della ghiandola endometriale;
Fattore di gravidanza precoce dell'uovo;
Proteine immunobloccanti del sinciziotrofoblasto;
HCG e lattogeno placentare (PL);
Licoproteine fibrinoidi placentari;
Proprietà proteolitiche del trofoblasto.
Frammentazione dell'embrione umano inizia alla fine del 1° giorno e continua per 3-4 giorni dopo la fecondazione (mentre l'embrione si sposta verso l'utero). La frammentazione dello zigote umano è completa, irregolare, asincrona. Durante il primo giorno avviene lentamente. La prima divisione viene completata dopo 30 ore; in questo caso il solco di clivaggio passa lungo il meridiano e si formano due blastomeri. Lo stadio dei due blastomeri è seguito dallo stadio dei quattro blastomeri. Dopo 40 ore si formano quattro cellule (Fig. 14, vedere riquadro).
Fin dalle prime divisioni si formano due tipi di blastomeri: “scuri” e “chiari”. I blastomeri “chiari” si frammentano più velocemente e si trovano in uno strato attorno a quelli “scuri”, che finiscono nel mezzo dell'embrione. Dai blastomeri “leggeri” superficiali nasce successivamente un trofoblasto, che collega l'embrione con l'organismo materno e gli fornisce nutrimento. I blastomeri "scuri" interni formano l'embrioblasto: da esso si formano il corpo dell'embrione e tutti gli altri organi extraembrionali, ad eccezione del trofoblasto. Quando la blastocisti entra nell'utero, aumenta di dimensioni a causa dell'aumento del numero di blastomeri e del volume del fluido dovuto al maggiore assorbimento delle secrezioni delle ghiandole uterine da parte del trofoblasto e alla produzione attiva di fluido da parte del trofoblasto stesso.
Nel trofoblasto aumenta il numero di lisosomi, in cui si accumulano enzimi che forniscono la lisi del tessuto uterino e quindi facilitano l'introduzione dell'embrione nello spessore della mucosa uterina, ad es. annidamento. L'impianto (nidazione) inizia il 7° giorno dopo la fecondazione e dura circa 40 ore (Fig. 15, vedere riquadro). In questo caso, la blastocisti è completamente circondata dal tessuto endometriale, la decidua.
Lo strato del trofoblasto si differenzia presto nello strato esterno: il sinciziotrofoblasto, che viene costantemente rifornito di nuclei e citoplasma a causa dello strato interno sottostante del citotrofoblasto (strato di Langhans), poiché la divisione nucleare si osserva solo nel citotrofoblasto. Il terzo derivato del trofoblasto non è proliferativo ed è un tipo di cellula mononucleare originariamente denominata "cellule X" ed è anche noto come "trofoblasto intermedio". Questo è il tipo principale di cellule che compongono la piattaforma placentare e, insieme alle cellule della decidua, penetrano nelle arterie spirali materne e formano anche la maggior parte delle cellule dei setti placentari. Le cellule X sono la principale fonte di lattogeno placentare umano (HPL - lattogeno placentare umano) e grandi quantità di proteine essenziali della gravidanza (MBP - proteina di base principale)
Durante le prime 2 settimane, il trofoblasto consuma i prodotti di degradazione dei tessuti materni (tipo di nutrizione istiotrofica). Quindi il sinciziotrofoblasto, crescendo sotto forma di villi e producendo enzimi proteolitici, penetra nell'utero, distrugge i vasi decidui materni, consentendo così al sangue della madre di riversarsi in lacune irregolari - che sono il futuro "spazio intervilloso". Pertanto, il trofoblasto entra in contatto diretto con il sangue dei vasi materni e l'embrione inizia a ricevere nutrimento direttamente dal sangue materno (nutrizione di tipo ematotrofo). La piena circolazione sanguigna nel feto viene stabilita approssimativamente nella quinta settimana dopo la fecondazione.
Una volta completato l'impianto, nello sviluppo dell'embrione inizia un periodo molto importante di organogenesi e placentazione. Dal 20° al 21° giorno, il corpo dell'embrione si separa dagli organi extraembrionali e avviene la formazione finale dei primordi assiali. L'organogenesi si completa entro la 12-16a settimana di vita intrauterina.
I periodi dello sviluppo prenatale sono mostrati in Fig. 16.
La massa embrionale si differenzia, si formano gli strati germinali: 1) ectoderma; 2) mesoderma; 3) endoderma. Sono anche differenziati (Fig. 17, vedere riquadro).
Il tubo neurale è formato dall'ectoderma. La chiusura del tubo neurale inizia nella regione cervicale, per poi diffondersi posteriormente e cranialmente, dove si formano le vescicole midollari. Approssimativamente il 25° giorno, il tubo neurale è completamente chiuso e dall'ambiente esterno
 Riso. 16. Periodi di sviluppo prenatale
Riso. 16. Periodi di sviluppo prenatale
Solo due aperture non chiuse comunicano tra loro alle estremità anteriore e posteriore: i neuropori anteriore e posteriore. Dopo altri 5-6 giorni, entrambi i neuropori crescono troppo. Quando le pareti laterali delle pieghe neurali si chiudono e si forma il tubo neurale, appare la cosiddetta cresta neurale. Le cellule della cresta neurale sono in grado di migrare. Nel tronco, le cellule migranti formano i gangli parasimpatici e simpatici e la midollare del surrene. Alcune cellule rimangono nella regione della cresta neurale, vengono segmentate e danno origine ai gangli spinali.
Differenziazione mesoderma inizia il 20° giorno dell'embriogenesi.
Le cellule del mesoderma si precipitano verso la superficie interna della cavità della blastocisti e si differenziano nel tessuto connettivo del corion e dei villi. Il luogo in cui queste cellule lasciano l'embrione diventa il cordone ombelicale, nel quale crescono i vasi allantoici della futura placenta.
I cambiamenti nell'embrione stesso sono espressi nel fatto che le sezioni dorsali dei fogli mesodermici sono divise in segmenti densi che giacciono sui lati della notocorda - somiti. Il processo di formazione dei segmenti, o somiti, inizia nella parte testa dell'embrione e si diffonde in direzione caudale. E se al 22esimo giorno di sviluppo l'embrione ha 7 paia di segmenti, al 35esimo giorno ci sono 44 paia. Nel processo di differenziazione del mesoderma, un rudimento nefrogenico ed embrionale
Il primo rudimento del tessuto connettivo è il mesenchima. Le cellule ecto ed endodermiche partecipano parzialmente alla formazione del mesenchima.
Endoderma forma una cavità: l'intestino primario, il futuro tubo digestivo, che si sviluppa attraverso lo stadio di formazione del sacco vitellino. La separazione dell'endoderma intestinale inizia con la comparsa della piega del tronco che, andando più in profondità, separa l'endoderma embrionale - l'intestino primario - dall'endoderma extraembrionale - il sacco vitellino. All'inizio della 4a settimana si forma un'invaginazione ectodermica all'estremità anteriore dell'embrione: la fossa orale. Approfondendosi, la fossa raggiunge l'estremità anteriore dell'intestino e, dopo aver sfondato la membrana che li separa, si trasforma nell'apertura orale del nascituro.
Il sacco vitellino e il tubo digerente rimangono collegati per qualche tempo attraverso il dotto onfalomesenterico (gambo vitellino), che termina in un potenziale diverticolo di Meckel. Il gambo del tuorlo, come il sacco vitellino, successivamente si atrofizza.
Pertanto, il sacco vitellino, formato dall'endoderma extraembrionale e dal mesoderma extraembrionale, prende parte attiva alla nutrizione e alla respirazione dell'embrione umano per un tempo molto breve. Il ruolo principale del sacco vitellino è ematopoietico. Come organo emopoietico, funziona fino alla 7-8a settimana, quindi subisce uno sviluppo inverso. Nella parete del sacco vitellino si formano le cellule germinali primarie - i gonoblasti, che migrano da esso con il sangue nei rudimenti delle gonadi.
Nella parte posteriore dell'embrione, l'intestino risultante comprende anche quella porzione dell'endoderma da cui origina l'escrescenza endodermica dell'allantoide.
L'allantoide è un piccolo processo a forma di dito dell'endoderma che cresce nel gambo amniotico. Nell'uomo l'allantoide non è molto sviluppato, ma la sua importanza nel garantire la nutrizione e la respirazione dell'embrione è ancora grande, poiché lungo di esso crescono i vasi verso il corion, i cui rami finali si trovano nello stroma dei villi. Al 2° mese di embriogenesi l'allantoide è ridotto.
Nella fig. 18 (vedi riquadro) mostra come appare un embrione a 4-5 settimane.
Durante i periodi di organogenesi (Fig. 19) e placentazione, a seguito dell'azione patogena di fattori ambientali nell'embrione e nel feto, vengono colpiti principalmente quegli organi e sistemi che si trovano in questo stato.
tempo nel processo di differenziazione. Per i vari anlage di organi embrionali, i periodi critici non coincidono nel tempo tra loro. Pertanto, l'azione di un fattore dannoso provoca solitamente deformità di vari organi e sistemi. La fase più sensibile dello sviluppo sono le prime 3-6 settimane di ontogenesi (il secondo periodo critico di sviluppo).
 Riso. 19. Periodi di organogenesi
Riso. 19. Periodi di organogenesi
CRESCITA INTRAuterina DEL FETALE
La dinamica della crescita fetale nell'utero è assicurata dall'interazione del potenziale genetico di ogni singolo feto e dell'ambiente intrauterino, che è principalmente associato alla funzione della placenta e all'omeostasi materna. La dinamica della crescita fetale durante la gravidanza fisiologica corrisponde all'età gestazionale (Tabella 1).
Tabella 1
Dinamica della crescita fetale
Dopo 27 settimane di gravidanza, la dinamica della crescita è influenzata dal sesso del feto (Tabella 2).
Tabella 2a
Settimane di gestazione | Centili di massa per ragazzi, g (A.V. Mazurin, I.M. Vorontsov, 2000) |
||||
Tabella 2b
Dinamica della crescita fetale a seconda del sesso
Settimane di gestazione | Centili di massa femminile, g (A.V. Mazurin, I.M. Vorontsov, 2000) |
||||
La discrepanza tra le dimensioni del feto e l'effettivo periodo di gravidanza è definita dal concetto di “ritardo della crescita intrauterina” (IUGR) del feto. Il criterio internazionale per l'IUGR è il peso e/o l'altezza del feto inferiore al normale per l'età gestazionale (10° centile e inferiore). La sindrome IUGR è una delle manifestazioni cliniche dell'insufficienza placentare.
Placenta- un organo extraembrionale attraverso il quale si stabilisce una connessione tra l'embrione e il corpo della madre. La placenta umana appartiene al tipo di placenta villosa emocoriale discoidale. La formazione della placenta inizia alla 3a settimana, quando i vasi iniziano a crescere nei villi secondari (epiteliomesenchimali), formando i villi terziari, e termina alla 14a-16a settimana di gravidanza.
La placenta è divisa nella parte embrionale, o fetale, e nella parte materna, o uterina.
La parte fetale è rappresentata da un corion ramificato e dalla membrana amniotica ad esso attaccata, e la parte materna è rappresentata da una parte basale modificata dell'endometrio.
La parte embrionale, o fetale, della placenta entro la fine del 3o mese è rappresentata da una placca corionica ramificata, costituita da tessuto connettivo fibroso (collagene) ricoperto di cito e sinciziotrofoblasto. I villi ramificati del corion (gambo, o ancora, villi) sono ben sviluppati solo sul lato rivolto
miometrio. Qui attraversano l'intero spessore della placenta e con i loro apici si immergono nella parte basale dell'endometrio distrutto.
L'unità strutturale e funzionale della placenta formata è il cotiledone, formato dai villi del fusto e dai suoi rami secondari e terziari. Il numero totale di cotiledoni nella placenta raggiunge i 200.
La parte materna della placenta è rappresentata dalla placca basale e dai setti del tessuto connettivo che separano i cotiledoni gli uni dagli altri, nonché dalle lacune riempite di sangue materno.
Sulla superficie della placca basale, di fronte ai villi coriali, è presente una sostanza amorfa: il fibrinoide di Rohr. Le cellule trofoblastiche della lamina basale, insieme ai fibrinoidi, svolgono un ruolo significativo nel garantire l'omeostasi immunologica nel sistema madre-feto.
Il sangue nelle lacune si rinnova continuamente. Proviene dalle arterie uterine, che entrano qui dal rivestimento muscolare dell'utero. Queste arterie corrono lungo i setti placentari e si aprono in lacune. Il sangue materno scorre dalla placenta attraverso le vene che originano dalle lacune.
Il sangue della madre e il sangue del feto circolano attraverso sistemi vascolari indipendenti e non si mescolano tra loro. La barriera emocoriale che separa entrambi i flussi sanguigni è costituita dall'endotelio dei vasi fetali, dai vasi circostanti del tessuto connettivo, dall'epitelio dei villi coriali (citotrofoblasto, sinciziotrofoblasto) e inoltre dal fibrinoide, che in alcuni punti copre i villi dall'esterno .
La placenta svolge funzioni trofiche, escretorie (per il feto), endocrine (produce hCG, progesterone, PL, estrogeni, ecc.), protettive (compresa la protezione immunologica).
Valore HCG
Stimola la produzione di progesterone da parte del corpo luteo.
Stimola le cellule di Leydig dei feti maschi e la produzione di testosterone.
Determina lo sviluppo degli organi genitali maschili.
È un indicatore precoce della gravidanza.
È un criterio per valutare l'efficacia del trattamento dei tumori trofoblastici, nonché un induttore dell'ovulazione per la sua somiglianza biologica con PH
Proprietà PL
Partecipa alla difesa immunologica - inibisce i linfociti materni.
Stimola la lipolisi e aumenta la concentrazione di acidi grassi liberi.
Inibisce la gluconeogenesi materna.
Aumenta i livelli plasmatici di insulina.
Stimola la sintesi di proteine e aminoacidi grazie all'effetto insulinogenico.
La concentrazione di PL dipende dal peso della placenta.
Membrana amniotica.È avascolare e costituisce la parete più interna del ricettacolo del frutto. La sua funzione principale è la produzione di liquido amniotico, che fornisce un ambiente per l'organismo in via di sviluppo e lo protegge dai danni meccanici. L'epitelio dell'amnio, rivolto verso la sua cavità, secerne il liquido amniotico e partecipa anche al loro riassorbimento. In questo caso, nell'epitelio dell'amnio che copre il disco placentare, avviene prevalentemente la secrezione, e nell'epitelio dell'amnio extraplacentare avviene prevalentemente il riassorbimento del liquido amniotico. Il liquido amniotico crea l'ambiente acquoso necessario per lo sviluppo dell'embrione, mantenendo la composizione e la concentrazione necessarie di sali nel liquido amniotico fino alla fine della gravidanza. L'amnio svolge anche una funzione protettiva, impedendo l'ingresso di agenti nocivi nel feto.
L'amnio è vagamente connesso al corion, nel quale si trovano i vasi fetali. Il suo attaccamento al corion avviene intorno alla 12a settimana di gravidanza; Prima di questo, c'è uno spazio pieno di liquido tra l'amnio e il corion. Inoltre, l'amnio si muove spesso durante la gravidanza e può staccarsi anche molto prima della nascita. A volte forma anche dei cordoni che, se entrano in contatto con il feto, possono causare amputazioni prenatali e altre deformità. Poiché l'amnio è collegato al cordone ombelicale ed è strettamente attaccato ad esso, i resti dei cordoni si trovano più spesso nel sito di attacco del cordone ombelicale.
Cordone ombelicale
Il cordone ombelicale è formato principalmente dal mesenchima situato nel peduncolo amniotico e nel peduncolo vitellino. Alla formazione del cordone prendono parte anche l'allantoide e i vasi che crescono lungo di esso. In superficie, tutte queste formazioni sono circondate dalla membrana amniotica. Il gambo del tuorlo e l'allantoide si riducono rapidamente e solo i loro resti si trovano nel cordone ombelicale del neonato.
Il cordone ombelicale formato è una formazione elastica di tessuto connettivo in cui passano due arterie ombelicali e una vena ombelicale. È formato da un tipico tessuto gelatinoso (mucoso), che contiene un'enorme quantità di acido ialuronico. È questo tessuto, chiamato gelatina di Wharton, che fornisce il turgore e l'elasticità del cordone. Protegge i vasi ombelicali dalla compressione, garantendo così un apporto continuo di nutrienti e ossigeno all'embrione.
Normalmente il cordone ombelicale è attaccato al disco placentare (attacco centrale), nel 7% è presente un attacco marginale (battagliatore) e nell'1% - sulle membrane (attacco meccanico). Gli attaccamenti anormali sono più comuni nelle gravidanze multiple. L'impianto della placenta non è associato ad anomalie fetali, ma può essere pericoloso a causa dell'aumentata incidenza di trombosi vascolari e della possibilità di sanguinamento dai vasi rotti durante il parto.
La lunghezza del cordone ombelicale è in gran parte determinata dall'attività motoria del feto. Pertanto, un cordone ombelicale corto indica spesso la sua immobilità a causa di patologie neuromuscolari o fusioni amniotiche. Al contrario, un cordone ombelicale lungo è talvolta il risultato di una maggiore attività motoria del feto.
Una singola arteria del cordone ombelicale è presente in più dell'1% dei casi, più spesso nelle gravidanze multiple. Circa la metà di questi neonati presenta anomalie congenite, alcune delle quali dovrebbero essere diagnosticate attivamente, e altri problemi perinatali. Una sola arteria del cordone ombelicale, invece, può essere presente in un neonato del tutto normale; quindi questo riscontro segnala solo la necessità di cautela riguardo alla presenza di patologia in questo neonato.
Nonostante il fatto che il corpo della madre e del feto siano geneticamente estranei nella composizione delle proteine, di solito non si verifica un conflitto immunologico.
passeggiate. Ciò è garantito da una serie di fattori; Di questi, i seguenti sono particolarmente importanti:
1 - proteine sintetizzate dal sinciziotrofoblasto che inibiscono la risposta immunitaria del corpo materno;
2 - hCG e PL, situati in alte concentrazioni sulla superficie del sinciziotrofoblasto, partecipano all'inibizione dei linfociti materni;
3 - effetto immunomascherante delle glicoproteine del fibrinoide placentare, caricate, come i linfociti del sangue di lavaggio, negativamente;
4 - proprietà proteolitiche del trofoblasto, che contribuiscono anche all'inattivazione delle proteine estranee;
5 - liquido amniotico con anticorpi che bloccano gli antigeni A e B (contenuti nel sangue di una donna incinta) e impediscono loro di entrare nel sangue del feto in caso di gravidanza incompatibile.
Nel processo di formazione del sistema madre-feto, ci sono una serie di periodi critici che sono più significativi per stabilire l'interazione tra i due sistemi e per creare condizioni ottimali per lo sviluppo fetale.
Nell'ontogenesi umana si possono distinguere diversi periodi critici dello sviluppo: nella progenesi, nell'embriogenesi e nella vita postnatale. Questi includono:
1) sviluppo delle cellule germinali - oogenesi e spermatogenesi;
2) fecondazione;
3) impianto (7-8 giorni di embriogenesi);
4) sviluppo dei primordi degli organi assiali e formazione della placenta (3-8a settimana di sviluppo);
5) stadio di maggiore crescita del cervello (15-20a settimana);
6) formazione dei principali sistemi funzionali del corpo e differenziazione dell'apparato riproduttivo (20-24a settimana);
7) nascita;
8) periodo neonatale (fino a 1 anno);
9) pubertà (11-16 anni).
Il ciclo ovarico è costituito da due fasi: follicolare e luteale, separate dall'ovulazione e dalle mestruazioni. La durata del ciclo ovarico (mestruale) varia normalmente dai 21 ai 35 giorni.
IN fase follicolare sotto l'influenza dell'FSH vengono stimolati la crescita e lo sviluppo di uno o più follicoli primordiali, nonché la differenziazione e la proliferazione delle cellule della granulosa. L'FSH stimola anche i processi di crescita e sviluppo dei follicoli primari, la produzione di estrogeni da parte delle cellule epiteliali follicolari. L'estradiolo, a sua volta, aumenta la sensibilità delle cellule della granulosa all'azione dell'FSH. Insieme agli estrogeni vengono secrete piccole quantità di progesterone. Dei tanti follicoli che iniziano a crescere, solo 1 raggiungerà la maturità finale, meno spesso 2-3. Il rilascio preovulatorio delle gonadotropine determina il processo stesso dell'ovulazione. Il volume del follicolo aumenta rapidamente parallelamente all'assottigliamento della parete follicolare. L'aumento significativo dei livelli di estrogeni osservato entro 2-3 giorni prima dell'ovulazione è dovuto alla morte di un gran numero di follicoli maturi con rilascio di liquido follicolare. Alte concentrazioni di estrogeni inibiscono la secrezione di FSH da parte della ghiandola pituitaria attraverso un meccanismo di feedback negativo. L'aumento ovulatorio di LH e, in misura minore, di FSH è associato all'esistenza di un meccanismo di feedback positivo di concentrazioni ultra elevate di livelli di estrogeni e LH, nonché a un forte calo dei livelli di estradiolo durante le 24 ore precedenti l'ovulazione .
Ovulazione dell'uovo si verifica solo in presenza di LH o gonadotropina corionica umana. Inoltre, FSH e LH agiscono come sinergici durante lo sviluppo del follicolo e in questo momento le cellule della teca secernono attivamente estrogeni.
Dopo l'ovulazione si verifica una forte diminuzione dei livelli di LH e FSH nel siero del sangue. A partire dal 12° giorno della seconda fase del ciclo si osserva un aumento di 2-3 giorni del livello di FSH nel sangue, che avvia la maturazione di un nuovo follicolo, mentre la concentrazione di LH durante tutta la seconda fase del ciclo il ciclo tende a diminuire.
La cavità del follicolo ovulato collassa e le sue pareti si raccolgono in pieghe. A causa della rottura dei vasi sanguigni al momento dell'ovulazione, si verifica un'emorragia nella cavità del follicolo postovulatorio. Una cicatrice del tessuto connettivo appare al centro del futuro corpo luteo - stigma
Il picco ovulatorio di LH e il successivo mantenimento di livelli elevati dell'ormone per 5-7 giorni attiva il processo di proliferazione e metamorfosi ghiandolare delle cellule della zona granulare con la formazione delle cellule luteiniche, cioè delle cellule luteiniche. arriva fase luteale del ciclo ovarico.
Le cellule epiteliali dello strato granulare del follicolo si moltiplicano intensamente e, accumulando lipocromi, si trasformano in cellule luteali; la membrana stessa è abbondantemente vascolarizzata. Lo stadio di vascolarizzazione è caratterizzato dalla rapida proliferazione delle cellule epiteliali della granulosa e dall'intensa crescita di capillari tra di loro. I vasi penetrano lateralmente nella cavità del follicolo postvulatorio teche interne nel tessuto luteinico in direzione radiale. Ogni cellula del corpo luteo è riccamente fornita di capillari. Il tessuto connettivo e i vasi sanguigni, raggiungendo la cavità centrale, la riempiono di sangue, avvolgono quest'ultimo, delimitandolo dallo strato di cellule luteiniche. Il corpo luteo ha uno dei più alti livelli di flusso sanguigno nel corpo umano. La formazione di questa rete unica di vasi sanguigni termina entro 3-4 giorni dopo l'ovulazione e coincide con il periodo di massimo splendore della funzione del corpo luteo (Bagavandoss P., 1991).
L'angiogenesi consiste di tre fasi: frammentazione della membrana basale esistente, migrazione delle cellule endoteliali e loro proliferazione in risposta ad uno stimolo mitogeno. L’attività angiogenica è controllata dai principali fattori di crescita: fattore di crescita dei fibroblasti (FGF), fattore di crescita epidermico (EGF), fattore di crescita derivato dalle piastrine (PLGF), fattore di crescita insulino-simile-1 (IGF-1), nonché citochine come come tumore del fattore necrotico (TNF) e interleuchine (IL-1; IL-6) (Bagavandoss P., 1991).
Da questo momento in poi il corpo luteo inizia a produrre quantità significative di progesterone. Il progesterone inattiva temporaneamente il meccanismo di feedback positivo e la secrezione delle gonadotropine è controllata solo dall'influenza negativa dell'estradiolo. Ciò porta ad una diminuzione del livello delle gonadotropine nella fase centrale del corpo luteo a valori minimi (Erickson G.F., 2000).
Il progesterone, sintetizzato dalle cellule del corpo luteo, inibisce la crescita e lo sviluppo di nuovi follicoli, partecipa anche alla preparazione dell'endometrio per l'impianto dell'ovulo fecondato, riduce l'eccitabilità del miometrio, sopprime l'effetto degli estrogeni su l'endometrio nella fase secretoria del ciclo, stimola lo sviluppo del tessuto deciduo e la crescita degli alveoli nelle ghiandole mammarie. Il plateau della concentrazione sierica di progesterone corrisponde al plateau della temperatura rettale (basale) (37,2-37,5 ° C), che è alla base di uno dei metodi per diagnosticare l'ovulazione avvenuta e costituisce un criterio per valutare l'utilità della fase luteinica. La base per l'aumento della temperatura basale c'è una diminuzione del flusso sanguigno periferico sotto l'influenza del progesterone, che riduce la perdita di calore. Un aumento del suo contenuto nel sangue coincide con un aumento della temperatura corporea basale, che è un indicatore dell'ovulazione.
Il progesterone, essendo un antagonista degli estrogeni, limita il loro effetto proliferativo nell'endometrio, nel miometrio e nell'epitelio vaginale, provocando la stimolazione della secrezione di glicogeno da parte delle ghiandole endometriali, riducendo lo stroma dello strato sottomucoso, ad es. provoca cambiamenti caratteristici nell'endometrio necessari per l'impianto di un ovulo fecondato. Il progesterone riduce il tono dei muscoli uterini e li fa rilassare. Inoltre, il progesterone provoca la proliferazione e lo sviluppo delle ghiandole mammarie e durante la gravidanza contribuisce all'inibizione del processo di ovulazione.Se la fecondazione non avviene, dopo 10-12 giorni il corpo luteo mestruale inizia a regredire, ma se l'ovulo fecondato ha penetrato nell'endometrio e la blastula risultante inizia a sintetizzare hCG, quindi diventa il corpo giallo corpo luteo della gravidanza.
Le cellule della granulosa del corpo luteo secernono l'ormone polipeptidico relaxina, che svolge un ruolo importante durante il parto, provocando il rilassamento dei legamenti pelvici e il rilassamento della cervice, inoltre aumenta la sintesi del glicogeno e la ritenzione idrica nel miometrio, riducendone la contrattilità.
Se la fecondazione dell'uovo non avviene, il corpo luteo entra nella fase di sviluppo inverso, che è accompagnato da mestruazioni. Le cellule luteali subiscono cambiamenti distrofici, diminuzione delle dimensioni e si osserva picnosi dei nuclei. Il tessuto connettivo, che cresce tra le cellule luteali in disintegrazione, le sostituisce e il corpo luteo si trasforma gradualmente in una formazione ialina: il corpo bianco.
Periodo di regressione del corpo luteo caratterizzato da una pronunciata diminuzione dei livelli di progesterone, estradiolo e inibina A. Una diminuzione dei livelli di inibina A ed estradiolo, nonché un aumento della frequenza degli impulsi di secrezione di Gn-RH assicurano la predominanza della secrezione di FSH su LH . In risposta ad un aumento dei livelli di FSH, si forma infine un pool di follicoli antrali, da cui in futuro verrà selezionato il follicolo dominante. La prostaglandina F 2 a, l'ossitocina, le citochine, la prolattina e i radicali 0 2 hanno un effetto luteolitico, che può essere la base per lo sviluppo dell'insufficienza del corpo luteo in presenza di un processo infiammatorio nelle appendici. Le mestruazioni si verificano sullo sfondo della regressione del corpo luteo. Alla fine, i livelli di estrogeni e progesterone raggiungono il minimo. In questo contesto, il centro tonico dell'ipotalamo e della ghiandola pituitaria viene attivato e aumenta la secrezione prevalentemente di FSH, che attiva la crescita dei follicoli. Un aumento del livello di estradiolo porta alla stimolazione dei processi proliferativi nello strato basale dell'endometrio, che garantisce un'adeguata rigenerazione dell'endometrio.
I cambiamenti ciclici dell'endometrio riguardano il suo strato superficiale, costituito da cellule epiteliali compatte, e lo strato intermedio, che viene rifiutato durante le mestruazioni.
Come è noto, si distingue tra la fase I - fase di proliferazione (fase precoce - 5-7 giorni, media - 8-10 giorni, tardiva - 10-14 giorni) e la fase II, fase di secrezione (precoce -15- 18 giorni - i primi segni di trasformazioni secretorie; media - 19-23 giorni, la secrezione più pronunciata; tardivo - 24-26 giorni, inizio regressione, regressione con ischemia - 26-27 giorni), fase III, fase sanguinamento o mestruazioni ( desquamazione - 28-2 giorni e rigenerazione - 3-4 giorni).
Bene la fase di proliferazione dura 14 giorni . I cambiamenti nell'endometrio che si verificano durante questa fase sono causati dall'azione di una quantità crescente di estrogeni secreti dal follicolo in crescita e maturazione (Khmelnitsky O.K., 2000).
Nella fase iniziale della fase di proliferazione(5-7 giorni del ciclo) l'endometrio è sottile, non c'è divisione dello strato funzionale in zone, la sua superficie è rivestita da epitelio cilindrico appiattito, di forma cubica. Le cripte ghiandolari hanno la forma di tubi diritti o leggermente contorti con un lume stretto; nelle sezioni trasversali hanno una forma rotonda o ovale. L'epitelio delle cripte ghiandolari è prismatico, i nuclei sono ovali, situati alla base, sono ben colorati, il bordo apicale delle cellule epiteliali al microscopio ottico appare liscio e ben definito.
Nella fase intermedia della fase di proliferazione l’attività della fosfatasi alcalina aumenta nell’endometrio. Nello stroma si osservano fenomeni di edema e allentamento. Il citoplasma delle cellule stromali diventa più distinguibile, i loro nuclei vengono rivelati abbastanza chiaramente e il numero di mitosi aumenta rispetto allo stadio iniziale. I vasi stromali sono ancora sporadici, con pareti sottili.
Nella fase finale della fase di proliferazione(11-14° giorno del ciclo) si nota un certo ispessimento dello strato funzionale, ma è ancora assente la suddivisione in zone. La superficie dell'endometrio è rivestita da un alto epitelio colonnare. Le strutture ghiandolari acquisiscono una forma più contorta, a cavatappi e sono più strettamente adiacenti tra loro rispetto agli stadi precedenti. L'epitelio delle cripte ghiandolari è alto cilindrico. I suoi bordi apicali appaiono lisci e chiari al microscopio ottico. La microscopia elettronica rivela i microvilli, che sono densi processi citoplasmatici ricoperti da una membrana plasmatica. Aumentando di dimensioni, creano un'area aggiuntiva per la distribuzione degli enzimi. È in questa fase che l'attività della fosfatasi alcalina raggiunge il suo massimo (Topchieva O.I. et al., 1978).
Al termine della fase di proliferazione l'esame ottico ottico rivela piccoli vacuoli subnucleari in cui si rilevano piccoli granuli di glicogeno. In questa fase, il glicogeno si forma in connessione con la secrezione preovulatoria dei gestageni nel follicolo che ha raggiunto la maturità. Le arterie spirali dello stroma, che si sviluppano dallo strato basale fino allo stadio intermedio della fase proliferativa, non sono ancora molto tortuose, pertanto nelle sezioni istologiche si riscontrano solo uno o due vasi con pareti sottili tagliate trasversalmente (Topchieva O.I. et al. , 1978; Zheleznov B. I., 1979).
Pertanto, gli estrogeni, contemporaneamente alla proliferazione delle cellule epiteliali, stimolano lo sviluppo dell’apparato secretorio della cellula durante la fase di proliferazione, preparandola per un’ulteriore piena funzione nella fase di secrezione. Ciò spiega la sequenza di eventi che ha un profondo significato biologico. Questo è il motivo per cui senza una precedente esposizione agli estrogeni sull’endometrio, il progesterone non ha praticamente alcun effetto. Oggi è stato scoperto che i recettori del progesterone, che forniscono sensibilità a questo ormone, vengono attivati dalla precedente azione degli estrogeni.
La fase di secrezione dura 14 giorni, direttamente correlato all'attività ormonale del corpo luteo e alla corrispondente secrezione di progesterone. Una riduzione o un prolungamento della fase di secrezione di oltre due giorni nelle donne in età riproduttiva dovrebbe essere considerata una condizione patologica, poiché tali cicli, di regola, sono anovulatori. All'inizio o alla fine del periodo riproduttivo possono verificarsi fluttuazioni della fase secretoria da 9 a 16 giorni, ad es. con la formazione o l’estinzione del ciclo utero-ovarico.
Nella diagnosi della 1a settimana della fase secretoria, i cambiamenti nell'epitelio sono di particolare importanza, permettendoci di parlare dell'ovulazione avvenuta. I cambiamenti caratteristici nell'epitelio durante la prima settimana sono associati all'aumento della funzione del corpo luteo. Nella 2a settimana, il giorno dell'ovulazione passata può essere determinato con maggiore precisione dallo stato delle cellule stromali. I cambiamenti nella 2a settimana nello stroma sono associati alla massima funzione del corpo luteo e alla sua successiva regressione e diminuzione della concentrazione di progesterone.
Durante la fase iniziale della fase di secrezione(al 15-18° giorno del ciclo) lo spessore dell'endometrio aumenta notevolmente rispetto alla fase di proliferazione. Il segno più caratteristico dell'inizio della fase di secrezione - la sua fase iniziale - è la comparsa di vacuoli subnucleari nell'epitelio delle ghiandole. In uno studio ottico-luce convenzionale, la manifestazione della secrezione sotto forma di vacuoli subnucleari si osserva solitamente il 16 ° giorno del ciclo, il che indica che si è verificata l'ovulazione e la pronunciata funzione ormonale del corpo luteo mestruale. Entro il 17° giorno del ciclo (3° giorno dopo l'ovulazione), i granuli di glicogeno sono contenuti nella maggior parte delle ghiandole e si trovano allo stesso livello nelle regioni basali delle cellule sotto il nucleo. Di conseguenza, anche i nuclei situati sopra i vacuoli sono disposti in fila, allo stesso livello. Quindi, il 18° giorno (4° giorno dopo l'ovulazione), i granuli di glicogeno si spostano nelle parti apicali delle cellule, come se bypassassero il nucleo. Di conseguenza, i nuclei sembrano discendere nuovamente fino alla base della cellula. Spesso a questo punto i nuclei di cellule diverse si trovano a livelli diversi. Anche la loro forma cambia: diventano più arrotondate, le mitosi scompaiono. Il citoplasma delle cellule diventa basofilo e nella loro parte apicale vengono rilevati mucopolisaccaridi acidi.
La presenza di vacuoli subnucleari è un segno di ovulazione avvenuta. Dobbiamo però ricordare che sono chiaramente visibili al microscopio ottico 36-48 ore dopo l'ovulazione. Va tenuto presente che i vacuoli subnucleari possono essere osservati anche in altre situazioni caratterizzate dall'azione del progesterone. Allo stesso tempo, però, non verranno rilevati allo stesso modo in tutte le ghiandole e la loro forma e dimensione saranno diverse. Pertanto, i vacuoli subnucleari si trovano spesso nelle singole ghiandole nel tessuto dell'endometrio “misto” ipoplastico e iperplastico.
Insieme alla vacuolizzazione subnucleare, lo stadio iniziale della fase di secrezione è caratterizzato da un cambiamento nella configurazione delle cripte ghiandolari: sono tortuose, espanse, uniformi e regolarmente localizzate nello stroma lasso, alquanto edematoso, che indica l'azione del progesterone su elementi stromali. Le arterie a spirale nella fase iniziale della fase di secrezione acquisiscono un aspetto più tortuoso, ma non si osservano ancora i “grovigli” caratteristici delle fasi successive di secrezione.
Nella fase intermedia della fase di secrezione(19-23 giorni del ciclo) nell'endometrio si osservano le trasformazioni secretorie più pronunciate, che si verificano a causa della più alta concentrazione di ormoni del corpo luteo. Lo strato funzionale è ispessito. Presenta chiaramente una divisione in strati spugnosi (spugnosi) o profondi e compatti o superficiali. Nello strato compatto le cripte ghiandolari sono meno tortuose, predominano le cellule stromali, l'epitelio che riveste la superficie dello strato compatto è alto, prismatico e non secernente. Le cripte ghiandolari a forma di cavatappi sono abbastanza strettamente adiacenti l'una all'altra, i loro lumi si espandono sempre più, soprattutto entro il 21-22esimo giorno del ciclo (cioè entro il 7-8o giorno dopo l'ovulazione) e diventano più piegate. Il processo di rilascio di glicogeno mediante secrezione apocrina nel lume delle ghiandole termina entro il 22° giorno del ciclo (8° giorno dopo l'ovulazione), che porta alla formazione di ghiandole grandi e allungate piene di granuli fini che sono chiaramente visibili quando colorati per glicogeno.
Nello stroma, durante la fase intermedia della fase di secrezione, si verifica una reazione di tipo deciduale, che si nota principalmente attorno ai vasi. Successivamente la reazione decidua da tipo insulare acquista carattere diffuso, soprattutto nelle parti superficiali dello strato compatto. Le cellule del tessuto connettivo diventano grandi, di forma rotonda o poligonale, ricordando l'aspetto di un pavimento terminale; l'ottavo giorno dopo l'ovulazione, in esse si trova il glicogeno.
L'indicatore più accurato della fase intermedia della secrezione, che indica un'alta concentrazione di progesterone, sono i cambiamenti nelle arterie a spirale, che nella fase intermedia della secrezione sono bruscamente tortuose e formano “grovigli”. Si trovano non solo nello strato spugnoso, ma anche nelle parti più superficiali dello strato compatto, poiché dal 9° giorno dopo l'ovulazione diminuisce l'edema stromale, poi dal 23° giorno del ciclo i grovigli delle arterie spirali sono già più chiaramente espresso. La presenza di vasi a spirale sviluppati nello strato funzionale dell'endometrio è considerata uno dei segni più affidabili che determinano il pieno effetto del progesterone. Il debole sviluppo dei “grovigli” dei vasi a spirale nell'endometrio della fase secretoria è considerato una manifestazione di una funzione insufficiente del corpo luteo e di un'insufficiente preparazione dell'endometrio all'impianto.
Come indicato da O.I. Topchieva et al. (1978), la struttura dell'endometrio della fase secretoria intermedia al 22-23 giorno del ciclo può essere osservata con una funzione ormonale prolungata e aumentata del corpo luteo mestruale, cioè con persistenza del corpo luteo (in questi casi, la succosità e la trasformazione deciduale dello stroma, nonché la funzione secretoria delle ghiandole, sono particolarmente pronunciate), o nelle prime fasi della gravidanza durante i primi giorni dopo l'impianto - con gravidanza intrauterina al di fuori della zona di impianto; così come uniformemente in tutte le parti della mucosa del corpo uterino con gravidanza ectopica progressiva.
Fase tardiva della fase di secrezione(24-27° giorno del ciclo) si verifica se la fecondazione dell'ovulo non è avvenuta e la gravidanza non è avvenuta. In questo caso, il 24 ° giorno del ciclo (10 ° giorno dopo l'ovulazione), il trofismo dell'endometrio, a causa dell'inizio della regressione del corpo luteo e, di conseguenza, una diminuzione della concentrazione di progesterone, viene interrotto e in esso si sviluppano numerosi processi distrofici, ad es. Cambiamenti regressivi si verificano nell'endometrio.
Con la microscopia ottica convenzionale, 3-4 giorni prima delle mestruazioni previste (il 24-25° giorno del ciclo), si nota una diminuzione della succosità dell'endometrio a causa della perdita di liquido e dell'increspatura dello stroma dell'endometrio. si osserva lo strato funzionale. A causa dell'increspamento dello stroma endometriale, le ghiandole diventano ancora più piegate, sono vicine l'una all'altra e acquisiscono una forma a dente di sega nelle sezioni longitudinali e un contorno a forma di stella nelle sezioni trasversali. Oltre alle ghiandole in cui la funzione secretoria è già cessata, c'è sempre un certo numero di ghiandole con una struttura corrispondente ai primi stadi della fase secretoria. L'epitelio delle cripte ghiandolari è caratterizzato da colorazione disomogenea dei nuclei, alcuni dei quali picnotici; nel citoplasma compaiono piccole gocce di lipidi.
Durante questo periodo, nello stroma, le cellule predeciduali si avvicinano l'una all'altra e vengono rilevate non solo sotto forma di isole attorno ai grovigli di vasi a spirale, ma anche diffusamente in tutto lo strato compatto. Tra le cellule predeciduali si trovano piccole cellule con nuclei scuri: cellule granulari endometriali che, come dimostrato da studi al microscopio elettronico, vengono trasformate da cellule del tessuto connettivo, ad es. cellule predeciduali più grandi, che si trovano prevalentemente in uno strato compatto. In questo caso, le cellule sono impoverite di glicogeno, i loro nuclei diventano picnotici.
Al 26-27° giorno del ciclo si possono riscontrare dilatazione dei capillari ed emorragie negli strati superficiali dello stroma. Questo perché man mano che il ciclo avanza, le arteriole spirali si allungano più velocemente di quanto aumenta lo spessore dell'endometrio, per cui i vasi si adattano all'endometrio aumentando la tortuosità. Durante il periodo premestruale, la spirale diventa così pronunciata da rallentare il flusso sanguigno e causare stasi e trombosi. Questo punto, insieme ad una serie di altri processi biochimici, spiega la necrosi endometriale e i cambiamenti distrofici nei vasi sanguigni che portano al sanguinamento mestruale. Poco prima dell'inizio delle mestruazioni, la vasodilatazione viene sostituita dallo spasmo, che si spiega con l'azione di vari tipi di prodotti tossici di degradazione proteica o di altre sostanze biologicamente attive sullo sfondo di un calo dei livelli di progesterone.
Fase di sanguinamento, mestruazioni(28-4° giorno del ciclo), caratterizzato da una combinazione di processi di desquamazione e rigenerazione.
Il sistema sensoriale del corpo.
118. Caratterizzare la struttura e le funzioni dell'organo della vista.
Risposta: comprende: il bulbo oculare e gli organi ausiliari dell'occhio (muscoli lisci della mela, palpebre, ciglia e paio lacrimale)
La struttura del bulbo oculare. Ha la forma di una cuccetta ed è costituito da gusci e un nucleo interno
Conchiglie: 1) Esterno (fibroso) il suo ruolo principale è protettivo. , e dà anche forma; le sue sezioni si distinguono: a) anteriore. Cornea. Trasparente, senza vasi, un numero enorme di terminazioni nervose. B) sclera. bianco, denso, ha vasi sanguigni, contiene terminazioni nervose.
2) Guscio medio. Tubazioni vascolari, scure, funzionali. Ci sono 3 sezioni: a) anteriore. L'iride (iride): al centro c'è un foro (pupilla), l'iride regola il flusso dei raggi luminosi nell'occhio modificando la dimensione della pupilla. Per fare questo, ci sono 2 muscoli nella pupilla: 1 - dilata la pupilla, 2 - restringe la pupilla. Questa sezione contiene pigmenti, la cui quantità determina il colore degli occhi. b) Sezione centrale. La coroide è una parte ispessita della membrana, situata sotto forma di un anello attorno al cristallino, che è attaccata al corpo ciliare con l'aiuto dei tendini. La base è costituita da tessuto connettivo lasso e contiene le cellule muscolari oculari che compongono il muscolo ciliare. Partecipa all'accomodamento dell'occhio (la capacità dell'occhio di vedere bene a diverse distanze) c) parte posteriore. Propria coroide dell'occhio, nutrizione del bulbo oculare.
3) Calotta interna. Retina. Colore giallastro, lamina oculare sottile, nessuna sensibilità alla luce (parte cieca), seconda sezione posteriore, fotosensibile. Contiene fino a 10 strati di cellule nervose. Importante primo 3:1) è costituito da cellule nervose visive o fotorecettrici. hanno forme diverse sotto forma di coni, bastoncelli, sono cellule nervose sensibili alla luce. I bastoncini formano sentimenti elevati. poiché funziona quando cambia l'illuminazione, danno la visione dei colori. Coni: sensi deboli. Funzionano in piena luce, ci sono 3 tipi di coni per la percezione del colore: 1) eritrolab 2) clorolab 3) cianolab. Una persona distingue circa 150 colori. Il secondo livello collega il livello 1 al livello 3. Il terzo strato, i neuroni di grandi dimensioni, formano il nervo ottico. Si forma proprio al centro della retina e forma i dischi ottici. Sul lato della macchia cieca ci sono diverse aree colorate di giallo, chiamate macchia gialla, e in questo punto ci sono i coni, ma non i bastoncelli.
Nucleo interno del bulbo oculare.
1) L'umor acqueo è un liquido che lava la cornea ed è coinvolto nel mantenimento della pressione intraoculare.
2) Lente - ha la forma di una lente convessa
3) Il corpo vitreo è una sostanza gelatinosa.
L'intero nucleo è trasparente e nel suo insieme costituisce il sistema ottico dell'occhio.
119. Caratterizzare la struttura e le funzioni dell'organo dell'udito e dell'equilibrio.
Risposta: Ci sono 3 dipartimenti:
1) Orecchio esterno. Include: padiglione auricolare e canale uditivo esterno.Funzione. catturare e condurre le vibrazioni sonore. Il padiglione auricolare è costituito da cartilagine, il padiglione auricolare esterno ha sporgenze educative: a) elica - il bordo della conca inferiore b) antelice - anteriore alla conca c) trago - una sporgenza limitata al meato uditivo. ti permettono di captare i suoni.
Canale uditivo esterno. Il canale curvo nell'adulto è lungo fino a 3,5 cm, inizia dall'apertura esterna e termina nel timpano cieco. Ci sono 2 sezioni: a) cartilaginea b) ossea (osso temporale) collegate tra loro ad angolo ottuso e aperte anteriormente e inferiormente. Quando si esamina il canale uditivo esterno, viene raddrizzato, spostando il padiglione auricolare indietro e verso l'alto. Nei bambini piccoli, queste 2 sezioni sono collegate. In modo leggermente diverso, la tirano indietro e verso il basso.
Timpano. Una sottile lamina trasparente forma un tessuto connettivo, con una depressione al centro all'esterno.
2) Orecchio medio. Comprende: cavità timpanica, 3 ossicini uditivi, tromba uditiva (di Eustachio). Parte dell'organo uditivo che conduce il suono. Situato nella piramide dell'osso temporale.
Cavità timpanica: ci sono 6 pareti, 5 delle quali sono formate dall'osso temporale e la sesta parete è formata internamente. Sulla superficie del timpano in questa cavità sono presenti 3 ossicini uditivi: 1) martello 2) incudine 3) staffa. Queste ossa sono collegate tra loro da articolazioni e + 2 muscoli: 1) tesi 2) indeboliti
Tromba di Eustachio: l'aria viene trasportata attraverso questo tubo per equalizzare la pressione sul timpano. Nell'adulto è lungo circa 4 cm, mentre nei bambini è più largo e più corto.
3) Orecchio interno. Situato nella piramide dell'osso temporale. È costituito da labirinti: 1) il labirinto osseo e in esso il labirinto membranoso. L'organo dell'equilibrio fa parte dell'orecchio interno, dei canali semicircolari (apparato vistibolare) ed è collegato anche all'orbita.
120. Caratterizzare la struttura e le funzioni della pelle e dei suoi derivati.
Risposta: costituisce la copertura generale del nostro corpo. L'organo più importante che svolge le seguenti funzioni: protezione, escrezione, termoregolazione, scambio gassoso, organo del tatto; sono presenti recettori in tutta l'area. La pelle umana è composta da 2 strati.
1) Strato superficiale. Epidermide. (epitelio squamoso non cheratinizzante)
2) Strato profondo. Il tessuto connesso, misto a fibre elastiche, è costituito da 2 strati: a) lo strato superiore denso (sporge sulla superficie della pelle) b) lo strato inferiore di grasso sottocutaneo. il tessuto è più sciolto, il fusto del capello si trova. La fibra grassa è coinvolta nella termoregolazione.
Derivati del cuoio
Capelli- appendici filiformi cheratinizzate della pelle con uno spessore di 0,005-0,6 mm e una lunghezza da diversi millimetri a 1,5 m, il loro colore, dimensione e distribuzione sono associati all'età, al sesso, alla razza e alla zona corporea. I peli ricoprono tutto il corpo, ad eccezione dei palmi delle mani, delle piante dei piedi, delle superfici laterali, palmari o plantari delle dita, del bordo vermiglio delle labbra, del glande, del clitoride e delle piccole labbra. Dei 2 milioni di peli presenti sul corpo umano. Circa 100.000 si trovano sul cuoio capelluto. Sono divisi in tre tipi:
lungo - spesso, lungo, pigmentato, che copre il cuoio capelluto e dopo la pubertà - il pube, le ascelle e negli uomini - anche i baffi, la barba e altre parti del corpo;
ispido: sopracciglia spesse, corte, pigmentate, che formano ciglia, che si trovano nel canale uditivo esterno e nel vestibolo della cavità nasale;
vellus - sottile, corto, incolore, che copre il resto del corpo (numericamente dominante); sotto l'influenza degli ormoni durante la pubertà, alcune parti del corpo possono trasformarsi in lunghe.
I peli sono costituiti da un fusto che sporge sopra la pelle e da una radice immersa in essa fino al livello del tessuto adiposo sottocutaneo (i peli vellus hanno meno profondità dei peli lunghi e ispidi). La radice è circondata da un follicolo pilifero, una formazione epiteliale cilindrica che sporge nel derma e nell'ipoderma e intrecciata con una borsa pilifera del tessuto connettivo. Vicino alla superficie dell'epidermide, il follicolo forma un'espansione - un imbuto, nel quale scorrono i condotti del sudore apocrino e delle ghiandole sebacee. All'estremità distale (profonda) del follicolo c'è anche un'estensione: il bulbo pilifero, nel quale cresce la papilla pilifera del tessuto connettivo con un gran numero di vasi sanguigni che forniscono nutrimento al bulbo.
Cellule epiteliali del bulbo servono come elementi cambiali (matrice) che assicurano la crescita dei capelli. Si dividono e, spostandosi, differenziano, formando (a seconda della loro posizione nel bulbo) cellule di diverso tipo. Queste cellule, subendo cheratinizzazione, partecipano alla formazione di varie parti del capello e della sua guaina interna della radice. Il bulbo contiene anche melanociti, che causano la pigmentazione dei capelli.
Midollo del pelo(formato dalle cellule della parte centrale del bulbo) si trova solo nei capelli lunghi e ispidi, è costituito da grandi cellule vacuolate, debolmente pigmentate, disposte a colonna di monete e contenenti granuli ossifili di tricoialina (precursore della sostanza cornea) nella citoplasma. Le cellule si cheratinizzano completamente solo a livello delle ghiandole sebacee, riempiendosi di cheratina molle.
Corteccia dei capelli(formato dalla parte centrale del bulbo) si trova attorno al midollo ed è costituito da cellule appiattite a forma di fuso, che diventano rapidamente cheratinizzate, riempiendosi di cheratina dura (meccanicamente e chimicamente stabile).
Cuticola dei capelli(formato dal bordo esterno della parte mediana del bulbo) circonda la corteccia ed è costituito da cellule che si trasformano in scaglie cornee contenenti cheratina dura, che si sovrappongono tra loro a mosaico con bordi sporgenti verso l'alto.
Vagina epiteliale interna(formato dalla parte periferica del bulbo) circonda la radice del pelo fino a livello dei dotti delle ghiandole sebacee, dove scompare. dentro comprende tre strati, chiaramente visibile solo in prossimità del bulbo e confluisce più in alto in un unico strato corneo (dall'interno verso l'esterno):
- la cuticola dell'epitelio interno della vagina è simile alla cuticola del capello, tuttavia le sue scaglie contengono cheratina, i loro bordi sporgenti sono diretti verso il basso e sono collegati alle scaglie della cuticola del capello;
- lo strato interno (contenente granuli) (Huxley) vicino al bulbo è formato da cellule cubiche (contenenti granuli di tricoialina), che, spostandosi verso l'alto, si riempiono di cheratina morbida e vengono distrutte;
- Lo strato esterno (pallido) (Henle) è formato da una fila di cellule cubiche leggere, riempite di cheratina morbida e distrutte.
Vagina epiteliale esternaè una continuazione dell'epidermide nel follicolo. Perde lo strato corneo a livello delle ghiandole sebacee e, assottigliandosi in 1-2 strati, si fonde con il bulbo.
Il muscolo che solleva i capelli è costituito da cellule muscolari lisce, un'estremità è intrecciata nel follicolo pilifero, l'altra nello strato papillare del derma. Quando si contrae, la radice del pelo obliqua assume una posizione più verticale e la pelle nell'area di inserzione del muscolo viene retratta (“pelle d'oca”).
Crescita dei capelli effettuato ad una velocità media di 0,35 mm al giorno e avviene ciclicamente e in modo asincrono nelle aree adiacenti della pelle (nell'uomo). Il ciclo è composto da tre fasi:
- le fasi di crescita attiva (anagen) comprendono l'allungamento del follicolo, l'attivazione della papilla e del bulbo (cellule della matrice e melanociti), la crescita della vagina interna e dei capelli stessi. La durata della fase anagen è determinata geneticamente e dura dai 2 ai 5 anni sul cuoio capelluto (in media 1000 giorni);
- fasi di cambiamenti regressivi (catagen) sono caratterizzate dalla cessazione della divisione cellulare della matrice, dalla scomparsa dei processi melanocitari, dall'ispessimento della parte terminale dei capelli con la formazione del cosiddetto “fiasco capillare”, dalla distruzione della vagina interna, e accorciamento del follicolo. Dura sul cuoio capelluto, in media 2-3 settimane;
- fase di riposo (telogen) - il bulbo è trattenuto in un follicolo accorciato, la sua rimozione avverrà solo in anagen; la riproduzione e la cheratinizzazione delle cellule epiteliali non si verificano. Dura circa 100 giorni.
Fattori che influenzano la crescita dei capelli: la crescita è favorita dagli androgeni nelle zone androgeno-dipendenti, ma sulla testa la inibiscono, e con un'esposizione prolungata possono provocare un'atrofia irreversibile del follicolo (che porta alla calvizie negli uomini geneticamente predisposti). Gli estrogeni rallentano la crescita e prolungano l'anagen, il cortisolo inibisce e la tiroxina accelera l'inizio dell'anagen.
Chiodoè una formazione sotto forma di placca che giace sulla superficie dorsale della falange distale delle dita. È costituito dalla lamina ungueale e dal letto ungueale.
Piastra per unghieè costituito da cheratina dura, è formato da numerosi strati di scaglie cornee, saldamente collegate tra loro, e giace sul letto ungueale. La sua parte prossimale è la radice dell'unghia, situata nella fessura ungueale posteriore e coperta eponichio(cuticola), ad eccezione di una piccola zona chiara a forma semilunare (luna). Distalmente la placca termina con un bordo libero che giace sopra la placca subungueale (iponichia). Lateralmente, la lamina ungueale è limitata da due pieghe della pelle: le pieghe delle unghie, dalle quali è separata dalle fessure ungueali laterali.
Letto per unghie- una formazione epiteliale costituita da uno strato basale e spinoso e che si trova sotto la lamina ungueale, che corrisponde al suo strato corneo. Forma creste epidermiche longitudinali, alternate alle pieghe del derma sottostante, che contiene numerosi vasi, nonché fibre collagene ed elastiche, fissandolo saldamente al periostio delle ossa falangee.
Matrice dell'unghia- parte prossimale ispessita del letto ungueale, formata da cellule che si dividono attivamente (il suo bordo distale corrisponde al bordo del foro). Le cellule appena formate sono incluse nella radice dell'unghia, dove si trasformano rapidamente (senza formazione di cheratoialina) in squame cornee, garantendo un movimento lento e continuo della lamina ungueale lungo il letto, con una velocità media di 0,1 mm al giorno (su le mani), crescita delle unghie. Strisce luminose trasversali sulla lamina ungueale indicano una disfunzione temporanea della matrice, quelle longitudinali - sul suo danno locale.
Afflusso di sangue alla pelle caratterizzato dalla presenza di due plessi arteriosi e venosi: profondi e superficiali, collegati da vasi di collegamento. Plesso profondo formato da una rete di vasi nelle parti profonde del derma e del tessuto adiposo sottocutaneo. Plesso superficiale situate sotto lo strato papillare del derma, da esso le arteriole si dirigono nelle papille, dove si diramano in numerosi capillari con rivestimento endoteliale continuo che si riuniscono in venule. Capillari fenestrati si trovano nelle papille piliferi e circondano le ghiandole sudoripare. Sono presenti anastomosi arteriolo-venulari tipo glomico, fornendo la regolazione della temperatura cutanea (particolarmente numerosi nel derma delle dita, delle orecchie e del naso).
Innervazione della pelleè fornito da fibre nervose efferenti e afferenti che formano il plesso subepidermico e dermico. Le fibre efferenti innervano il tessuto muscolare liscio dei vasi sanguigni, il muscolo elevatore del pelo e le ghiandole sudoripare. Le fibre afferenti sono associate a terminazioni nervose incapsulate (corpi lamellari, fiaschi terminali, corpi tattili e altri), che giacciono nel derma e sono meccanorecettori, nonché a terminazioni libere nell'epidermide e nel derma (nocicettori e termorecettori).
In corso ciclo uterino gli ormoni ovarici prodotti nel follicolo e nel corpo luteo influenzano i cambiamenti ciclici del tono, dell'eccitabilità e dell'afflusso di sangue all'utero. Cambiamenti ciclici più significativi si verificano nell'endometrio. La loro essenza sta nel processo di proliferazione correttamente ripetuto, nel cambiamento qualitativo, nel rigetto e nel ripristino dello strato della mucosa, rivolto verso il lume dell'utero. Questo strato, che subisce cambiamenti ciclici, è chiamato strato funzionale dell'endometrio. Lo strato della mucosa adiacente allo strato muscolare dell'utero non subisce cambiamenti ciclici ed è chiamato strato basale.
Il ciclo uterino, come quello ovarico, dura 28 giorni (meno spesso 21 o 30-35 giorni). Consiste in: fase di desquamazione, fase di rigenerazione, fase di proliferazione e fase di secrezione.
Fase di desquamazione si manifesta con sanguinamento che dura 3-5 giorni (mestruazioni). Lo strato funzionale della mucosa, sotto l'influenza degli enzimi, si disintegra, viene rifiutato e viene rilasciato insieme al contenuto delle ghiandole uterine e al sangue dai vasi rotti. La fase della desquamazione endometriale coincide con l'inizio della morte del corpo luteo nell'ovaio.
Fase di rigenerazione la mucosa inizia durante il periodo di desquamazione e termina 5-6 giorni dall'inizio delle mestruazioni. Il ripristino dello strato funzionale della mucosa avviene a causa della proliferazione dell'epitelio dei resti delle ghiandole situate nello strato basale e attraverso la proliferazione di altri elementi di questo strato (stroma, vasi, nervi). La rigenerazione è dovuta all'influenza formata nel follicolo, il cui sviluppo inizia dopo la morte del corpo luteo.
Fase di proliferazione l'endometrio coincide con la maturazione del follicolo nell'ovaio e prosegue fino al 14° giorno del ciclo (con un ciclo di 21 giorni fino a 10-11 giorni). Sotto l'influenza dell'ormone estrogeno, che influenza gli elementi nervosi e i processi metabolici nell'utero, si verifica la proliferazione o la crescita dello stroma e la crescita della mucosa. Le ghiandole si allungano in lunghezza, poi si torcono come un cavatappi, ma non contengono secrezione. La mucosa dell'utero durante questo periodo si ispessisce 4-5 volte.
Fase di secrezione coincide con lo sviluppo del corpo luteo nell'ovaio e dura dal 14°-15° al 28° giorno, cioè fino alla fine del ciclo.
Sotto l'influenza dell'ormone del corpo luteo si verificano importanti trasformazioni qualitative nella mucosa uterina. Le ghiandole producono secrezioni, la loro cavità si espande e nelle pareti si formano sporgenze a forma di baia. Le cellule stromali si allargano e diventano leggermente arrotondate, somigliando alle cellule decidue prodotte durante la gravidanza. Glicogeno, fosforo, calcio e altre sostanze si depositano nella mucosa.
Come risultato di questi cambiamenti nella mucosa, si creano condizioni favorevoli per lo sviluppo dell'embrione in caso di fecondazione. Alla fine della fase di secrezione si nota la permeazione sierosa dello stroma e appare una diffusa infiltrazione leucocitaria dello strato funzionale. I vasi di questo strato si allungano, acquisiscono una forma a spirale, in essi si formano delle dilatazioni e il numero delle anastomosi aumenta.
L'endometrio è costituito da due strati: funzionale e basale. Lo strato funzionale cambia struttura sotto l'influenza degli ormoni sessuali e, se la gravidanza non si verifica, viene rifiutato durante le mestruazioni.
Fase proliferativa
L'inizio del ciclo mestruale è considerato il 1° giorno delle mestruazioni. Alla fine delle mestruazioni, lo spessore dell'endometrio è di 1-2 mm. L'endometrio è costituito quasi esclusivamente dallo strato basale. Le ghiandole sono strette, diritte e corte, rivestite da un epitelio colonnare basso, il citoplasma delle cellule stromali è quasi lo stesso.
Quando i livelli di estradiolo aumentano, si forma uno strato funzionale: l'endometrio si prepara all'impianto dell'embrione. Le ghiandole si allungano e diventano contorte. Il numero di mitosi aumenta. Man mano che proliferano, l'altezza delle cellule epiteliali aumenta e l'epitelio stesso cambia da fila singola a fila multipla al momento dell'ovulazione. Lo stroma è rigonfio e allentato, con nuclei cellulari e volume citoplasmatico aumentati. I vasi sono moderatamente tortuosi.
Fase secretoria
Normalmente l'ovulazione avviene il 14° giorno del ciclo mestruale. La fase secretoria è caratterizzata da alti livelli di estrogeni e progesterone. Tuttavia, dopo l’ovulazione, il numero di recettori degli estrogeni nelle cellule endometriali diminuisce. La proliferazione endometriale viene gradualmente inibita, la sintesi del DNA diminuisce e il numero di mitosi diminuisce. Pertanto, il progesterone ha un effetto predominante sull'endometrio nella fase secretoria.
Nelle ghiandole endometriali compaiono vacuoli contenenti glicogeno, che vengono rilevati utilizzando la reazione PAS. Al 16° giorno del ciclo, questi vacuoli sono piuttosto grandi, presenti in tutte le cellule e situati sotto i nuclei. Il 17° giorno i nuclei, spinti da parte dai vacuoli, si trovano nella parte centrale della cellula. Il 18 ° giorno compaiono i vacuoli nella parte apicale e i nuclei nella parte basale delle cellule, il glicogeno inizia a essere rilasciato nel lume delle ghiandole mediante secrezione apocrina. Le migliori condizioni per l'impianto si creano il 6°-7° giorno dopo l'ovulazione, cioè il 20-21° giorno del ciclo, quando l'attività secretoria delle ghiandole è massima.
Il 21° giorno del ciclo inizia la reazione decidua dello stroma endometriale. Le arterie spirali sono nettamente tortuose; successivamente, a causa della diminuzione dell'edema stromale, sono chiaramente visibili. Innanzitutto compaiono le cellule decidue, che gradualmente formano grappoli. Il 24° giorno del ciclo, questi accumuli formano accoppiamenti eosinofili perivascolari. Il 25° giorno si formano isole di cellule decidue. Entro il 26° giorno del ciclo, la reazione decidua diventa massima. Circa due giorni prima delle mestruazioni, il numero di neutrofili che migrano lì dal sangue aumenta bruscamente nello stroma endometriale. L'infiltrazione dei neutrofili è sostituita dalla necrosi dello strato funzionale dell'endometrio.
Il ciclo mestruale uterino si verifica nell'utero, un ciclo di cambiamenti nell'endometrio.
Cambiamenti ciclici nell'endometrio riguardano il suo strato funzionale (superficiale), costituito da cellule epiteliali compatte, e lo strato intermedio, che viene rifiutato durante le mestruazioni.
Lo strato basale, che non viene rifiutato durante le mestruazioni, garantisce il ripristino degli strati desquamati.
In base ai cambiamenti dell'endometrio durante il ciclo si distinguono la fase di proliferazione, la fase di secrezione e la fase di sanguinamento (mestruazioni).
La trasformazione dell'endometrio avviene sotto l'influenza degli ormoni steroidei: la fase di proliferazione - sotto l'azione predominante degli estrogeni, la fase di secrezione - sotto l'influenza del progesterone e degli estrogeni.
Fase di proliferazione(follicolare) dura in media 12-14 giorni a partire dal 5° giorno del ciclo (Fig. 2.5). Durante questo periodo, si forma un nuovo strato superficiale con ghiandole tubolari allungate rivestite da un epitelio cilindrico con maggiore attività mitotica. Lo spessore dello strato funzionale dell'endometrio è di 8 mm.
Fase di secrezione (luteale) associato all'attività del corpo luteo, dura 14 giorni (±1 giorno) (Fig. 2.6). Durante questo periodo, l'epitelio delle ghiandole endometriali inizia a produrre secrezioni contenenti glicosaminoglicani acidi, glicoproteine e glicogeno.
L'attività della secrezione raggiunge il massimo tra il 20 e il 21 giorno. A questo punto, nell'endometrio viene rilevata la quantità massima di enzimi proteolitici e nello stroma si verificano trasformazioni decidue (le cellule dello strato compatto si allargano, acquisendo una forma rotonda o poligonale, il glicogeno si accumula nel loro citoplasma). Si nota una forte vascolarizzazione dello stroma: le arterie spirali sono nettamente tortuose, formando "grovigli" presenti in tutto lo strato funzionale. Le vene sono dilatate. Tali cambiamenti nell'endometrio, osservati nei giorni 20-22 (giorni 6-8 dopo l'ovulazione) del ciclo mestruale di 28 giorni, forniscono le migliori condizioni per l'impianto di un ovulo fecondato.
Entro il 24-27 giorno, a causa dell'inizio della regressione del corpo luteo e della diminuzione della concentrazione degli ormoni da esso prodotti, il trofismo endometriale viene interrotto con un graduale aumento dei cambiamenti degenerativi in esso. Dalle cellule granulari dello stroma endometriale vengono rilasciati granuli contenenti relaxina, che prepara il rigetto mestruale della mucosa. Nelle aree superficiali dello strato compatto si notano dilatazione lacunare dei capillari ed emorragie nello stroma, rilevabili 1 giorno prima dell'inizio delle mestruazioni.
Mestruazioni comprende la desquamazione e la rigenerazione dello strato funzionale dell'endometrio. A causa della regressione del corpo luteo e di una forte diminuzione del contenuto di steroidi sessuali nell'endometrio, aumenta l'ipossia. L'inizio delle mestruazioni è facilitato da uno spasmo prolungato delle arterie, che porta alla stasi del sangue e alla formazione di coaguli di sangue. L'ipossia tissutale (acidosi tissutale) è esacerbata dall'aumento della permeabilità dell'endotelio, dalla fragilità delle pareti dei vasi, da numerose piccole emorragie e dalla massiccia infiltrazione di leucociti. Gli enzimi proteolitici lisosomiali rilasciati dai leucociti migliorano la fusione degli elementi tissutali. A seguito di uno spasmo prolungato dei vasi, la loro espansione paretica avviene con aumento del flusso sanguigno. Allo stesso tempo, si verifica un aumento della pressione idrostatica nel microcircolo e la rottura delle pareti dei vasi sanguigni, che a questo punto hanno in gran parte perso la loro resistenza meccanica. In questo contesto si verifica la desquamazione attiva delle aree necrotiche dello strato funzionale. Entro la fine del 1° giorno delle mestruazioni, 2/3 dello strato funzionale vengono respinti e la sua completa desquamazione termina solitamente entro il 3° giorno.
La rigenerazione dell'endometrio inizia immediatamente dopo il rigetto dello strato funzionale necrotico. La base per la rigenerazione sono le cellule epiteliali dello stroma dello strato basale. In condizioni fisiologiche, già al 4° giorno del ciclo, l'intera superficie della ferita della mucosa risulta epitelizzata. Seguono nuovamente i cambiamenti ciclici nell'endometrio - le fasi di proliferazione e secrezione.
I cambiamenti consecutivi durante tutto il ciclo nell'endometrio - proliferazione, secrezione e mestruazioni - dipendono non solo dalle fluttuazioni cicliche dei livelli di steroidi sessuali nel sangue, ma anche dallo stato dei recettori tissutali per questi ormoni.
La concentrazione dei recettori nucleari dell'estradiolo aumenta fino alla metà del ciclo, raggiungendo il picco nell'ultimo periodo della fase di proliferazione endometriale. Dopo l'ovulazione si verifica una rapida diminuzione della concentrazione dei recettori nucleari dell'estradiolo, che continua fino alla fase secretoria tardiva, quando la loro espressione diventa significativamente inferiore rispetto all'inizio del ciclo.
La regolazione delle concentrazioni locali di estradiolo e progesterone è mediata in larga misura dalla comparsa di vari enzimi durante il ciclo mestruale. Il contenuto di estrogeni nell'endometrio dipende non solo dal loro livello nel sangue, ma anche dalla loro formazione nei tessuti. L'endometrio di una donna è in grado di sintetizzare
- In contatto con 0
- Google+ 0
- OK 0
- Facebook 0






