Campione
========================================
Cosso Cosso(L.)parafanghi anteriori farfalle dal grigio-marrone al grigio scuro con un motivo “marmo” e macchie grigio-bianche sfocate, nonché linee ondulate trasversali scure. Le ali posteriori sono marrone scuro con linee ondulate scure opache. Il petto è scuro superiormente, biancastro verso l'addome. L'addome scuro ha anelli chiari. Maschio ha un'apertura alare di 65-70 mm, femmina-da 80 a 95 mm. L'addome della femmina termina con un ovopositore retrattile e ben marcato. bruco subito dopo la schiusa rosso ciliegia, poi rosso carne. Testa e scudo occipitale di colore nero lucido. bruco adulto ha 8-11 cm (il più delle volte 8-9 cm), quindi è color carne giallastro, marrone in alto con una sfumatura viola. Lo scudo occipitale giallo-marrone presenta due macchie scure. Il foro di respirazione è marrone. Uovo ovale-longitudinale, bruno chiaro con striature nere, denso, di dimensioni 1,2 mm.
danni tiglio e bosco di pioppi, salici, querce, altre latifoglie e alberi da frutto. Le farfalle compaiono in natura dalla fine di giugno, soprattutto a luglio e, a seconda della posizione geografica, in alcune località anche prima di metà agosto. Volano lentamente in tarda serata. L'estate dura un massimo di 14 giorni. Durante il giorno si siedono in una posa caratteristica con il petto inclinato nella parte inferiore del tronco. Le femmine depongono le uova in gruppi di 15-50 pezzi nelle fessure della corteccia, in luoghi danneggiati, ferite cancerose dei tronchi ad altezze fino a 2 M. I bruchi si schiudono dopo 14 giorni. Innanzitutto, i tessuti liberiani vengono mangiati insieme. Sugli alberi più vecchi con corteccia spessa nella parte inferiore del tronco, i bruchi mangiano passaggi ovali lunghi, irregolari e irregolari nella sezione trasversale solo dopo il primo svernamento. Le pareti dei passaggi vengono distrutte da un liquido speciale e sono marroni o nere. Sui tronchi più sottili con corteccia liscia, i bruchi penetrano prima nel legno, di solito entro un mese dalla schiusa. I trucioli e gli escrementi del bruco vengono espulsi attraverso il foro inferiore. Alla fine della stagione di crescita, quando le foglie cadono, l'alimentazione dei bruchi si interrompe, che ibernano in passaggi fino alla fioritura del fogliame, cioè fino ad aprile - maggio, quando i bruchi continuano a nutrirsi nuovamente in passaggi separati fino all'autunno, ibernano una volta di più e finisci di mangiare. Si impupano o alla fine di un passaggio circolare, dove viene preparato in anticipo un foro per il volo, chiuso con schegge, o nel terreno, vicino a un tronco danneggiato in un bozzolo di schegge. La fase pupa dura 3-6 settimane. Prima di volare, la pupa, con l'aiuto delle spine, fuoriesce per metà dal foro di volo o dal bozzolo, in modo che la farfalla possa uscire più facilmente dall'esuvio. La generazione è di massimo due anni. esteso in tutta Europa, soprattutto al centro e al sud. È presente in tutta la zona forestale della parte europea della Russia, nel Caucaso, in Siberia e anche nell'Estremo Oriente. Conosciuto nella Cina occidentale e settentrionale e nell'Asia centrale.trascrizione
1 ACCADEMIA DELLE SCIENZE DELL'ISTITUTO ZOOLOGICO DELL'URSS CHIAVE DEGLI INSETTI DELLA PARTE EUROPEA DELL'URSS IN 5 VOLUMI II COLEOTTERI E FANOPTERI SOTTO L'EDIZIONE GENERALE DEL MEMBRO CORRISPONDENTE DELL'Accademia delle Scienze dell'URSS G. Ya. UKA" MOSCA LENINGRADO 1965
2 CHIAVI PER LA FAUNA DELL'URSS, PUBBLICATO DALL'ISTITUTO ZOOLOGICO DELL'Accademia delle Scienze dell'URSS, vol. 89 Compilato da: A. V. ALEKSEEV, L. V. ARNOLDI, E. L. GURYEVA, R. D. ZHANTIEV, V. A. ZASLAVSKY, J. D. KIRSHENBLAD, G. O. KRIVOLUTSKAYA, O. L. Kryzhanovsky, F. K. Lukyanovich, G. S. Medvedev, L. N. Medvedev, S. I. Medvedev , N. N. Plavilshchikov, A. A. Richter, V. A. Richter e M. E. Ter MINASYAN, D. S. SHAPIRO. Comitato editoriale: B. E. BYKHOVSKY, I. M. GROMOV, A. S. MONCHADSKY, O. A. SCARLATO, A. A. STRELKOV, A. A. SHTAKELBERG Curatori del volume E. L. GURYEVA e O. L. KRYZHANOVSKY
3 PREFAZIONE Il libro proposto costituisce il secondo volume della "Chiave degli insetti della parte europea dell'URSS" e comprende due ordini: un vasto complesso di coleotteri, o coleotteri (Coleoptera), e un piccolo ordine altamente specializzato di Fanwings ( Strepsitteri). Come nel volume I, la "Chiave" comprende tutti i generi di questi ordini conosciuti dalla parte europea dell'URSS e la stragrande maggioranza delle specie che vivono qui, comprese tutte le specie di importanza pratica. In generale sono state considerate più di 4000 specie appartenenti a 84 famiglie di Coleotteri e 4 famiglie di Fanotteri. Insieme a questo, alcune specie rare, scarsamente distribuite o scarsamente studiate non sono incluse nella "Chiave", e in un certo numero di famiglie che sono scarsamente studiate e non hanno alcun significato pratico evidente, le tabelle sono ridotte solo al genere. Tuttavia, i professionisti nel campo della protezione delle piante, della silvicoltura e dell'entomologia medica possono ottenere una risposta alla domanda sul nome e sulla distribuzione di tutte le specie importanti per l'agricoltura, la selvicoltura e la salute pubblica. Alcune parole e termini di uso frequente sono riportati sotto forma di abbreviazioni (per l'elenco delle abbreviazioni vedere p.??). Le specie nocive sono contrassegnate con un asterisco (*). Le misure del corpo sono espresse in millimetri, ma senza l'indicazione di "mm". I dati sul numero di specie conosciute dalla parte europea dell'URSS sono forniti per genere solo nei casi in cui non tutte le specie conosciute da qui sono incluse nella tabella chiave; la mancanza di dati sul numero delle specie indica che la tavola di questo genere comprende tutte le specie conosciute del territorio in esame. Le informazioni sulla biologia e sulla distribuzione geografica delle specie vengono fornite molto brevemente. Per le specie ampiamente distribuite, i dati sulla distribuzione vengono omessi; nei casi in cui vengono fornite queste informazioni, queste sono spesso limitate all'indicazione di grandi suddivisioni della parte europea dell'URSS (vedi mappa): nord, sud-est, centro, ecc. Il confine meridionale del territorio coperto dall'"Identificatore" corre lungo la costa del Mar Nero, compresa la costa settentrionale del Mar Caspio e il corso inferiore degli Urali; di conseguenza, la fauna dei versanti settentrionali della catena del Caucaso, che comprende un gran numero di specie specifiche caucasiche e mediterranee, solitamente non viene presa in considerazione. Solo nelle famiglie dei lamellari (Scarabaeidae), dei boscaioli (Cerambycidae), degli scarabei fogliari (Chrysomelidae) e dei punteruoli (Curculionidae) sono parzialmente incluse le specie caucasiche. Alla compilazione del volume II della Chiave hanno preso parte 17 entomologi che lavorano in numerosi istituti di ricerca scientifica e istituti di istruzione superiore. Volume I. Inferiore, alato antico, con metamorfosi incompleta. Ed. "Scienza", M. L., 1964, 936 pagine.
4 4 PREFAZIONE delle istituzioni educative del paese: L. V. Arnoldi, E. L. Guryeva, V. A. Zaslavsky, O. L. Kryzhanovsky, G. S. Medvedev, V. A. Richter, M. E. Ter Minasyan (Istituto zoologico dell'Accademia delle scienze dell'URSS), A. V. Alekseev (Istituto pedagogico, Orekhovo-Zuevo), R. D. Zhantiev (Università statale di Mosca), Ya. D. Kirshenblat (Istituto medico di Chernivtsi), G. O. Krivolutskaya (Sezione siberiana dell'Estremo Oriente dell'Accademia delle scienze dell'URSS, Vladivostok), L. N. Medvedev (Dipartimento di biologia generale dell'Università Accademia delle Scienze dell'URSS, Mosca), S. I. Medvedev e D. S. Shapiro (Università statale di Kharkov), Università N. N.). Inoltre, sono stati utilizzati i dati del defunto F. K. Lukyanovich e A. A. Richter (Istituto zoologico dell'Accademia delle scienze dell'URSS). La distribuzione delle singole parti dell'opera tra gli autori è indicata nelle relative sezioni del testo. Molti dei disegni sono stati realizzati dagli autori, la maggior parte dei ridisegni e dei disegni originali sono stati realizzati dall'artista S. S. Kozhevnikovyi, dall'artista V. N. Lyakhov per gli scarabei staliniani e dall'artista F. I. Gunyaev per gli scarabei pseudobark. I tipi di nuove forme descritti qui e nel volume I sono conservati presso l'Istituto Zoologico dell'Accademia delle Scienze dell'URSS a Leningrado. G. Ya. Bei-Bienko
6 6 COLEOTTERI O SCARABEO Fig. Fig. 1. Vista generale dello scarabeo saltatore (Cicindela) dall'alto con le ali destre spiegate e quelle sinistre rimosse (la parte superiore dell'ala destra non è mostrata). (Secondo Ganglbauer con modifiche). testa dell'obiettivo; MD uomo dibula; antenne baffi; palpi, palpi mascellari; labbro superiore vg; fascia di contanti; ch occhio; gr petto; prsp pronoto; scutello a scudo; hsp metanoto; elitra elitra; sutura sh; angolo di sutura sh; ala krl; br addome; spiracolo respiratorio; T1 T8 1 8° tergiti. Gli organi posti davanti al clipeo, alle guance e alla gola e collegati ad essi più o meno mobili sono detti organi orali, o parti orali; tutte le loro parti sono generalmente ben sviluppate. Il labbro superiore ha solitamente l'aspetto di una placca trasversale, talvolta retratta o fusa stabilmente con il clipeo, raramente (nei punteruoli e nei scolitidi) non sviluppata affatto. Ricopre più o meno dall'alto le mandibole (mascelle, o mandibole), che nella maggior parte dei casi sono ben sviluppate e fortemente chitinizzate, solo in alcuni scarabei stercorari sono in parte coriacee; all'interno portano spesso denti, e nei maschi di alcune specie (ad esempio nel cervo volante) raggiungono dimensioni enormi. Direttamente sotto di loro c'è una coppia di mascelle (mascelle inferiori, Fig. 3, 2), ciascuna delle quali è costituita da una piastra principale (cardo), uno stelo (gambo) e un palpo mascellare a 4 segmenti e due lobi masticatori attaccati al ultimo: interno (lacinia) ed esterno (galea); il lobo esterno nei coleotteri macinati e nelle famiglie vicine forma il secondo palpo mascellare. Dal basso, la mascella inferiore confina con il labbro inferiore (Fig. 3, 2), che consiste in un mento attaccato al margine anteriore del sottomento e solitamente separato da esso mediante una sutura; il mento sul bordo anteriore è spesso dentellato o reca una punta; davanti al mento è attaccata una lingua, a quest'ultimo sono attaccati 2 palpi labiali trisegmentati e 2 lingue accessorie (paraglossi). Baffi e k e (antenne) si trovano sulla fronte o sulle guance; il luogo del loro attacco è chiamato cavità antennale; sono generalmente costituiti da 11 segmenti, ma il numero dei segmenti può talvolta diminuire (fondendosi) talvolta fino a 2 (Paussus, Claviger) oppure dividendo i singoli segmenti può aumentare fino a 40 o più . La struttura delle antenne nei coleotteri è estremamente varia ed è di grande importanza nella determinazione. Le antenne filiformi (Fig. 4, 1) sono costituite da segmenti cilindrici e hanno all'incirca lo stesso spessore su tutta la sua lunghezza (solo il 1° segmento è spesso più spesso o più lungo degli altri). Le antenne setolate (Fig. 4, 2) differiscono da quelle filiformi in quanto si assottigliano gradualmente verso l'apice. Antenne a perline (Fig. 4, 3) è costituito da brevi segmenti arrotondati separati da costrizioni. Le antenne seghettate (seghettate) (Fig. 4, 4) hanno segmenti triangolari diretti ad angoli acuti su un lato. Nelle antenne a forma di pettine (Fig. 4, 5), questi angoli sono allungati in lunghi processi. Le antenne debolmente e gradualmente allargate verso l'apice sono dette ispessite (Fig. 4, 6). Le antenne a forma di clava (Fig. 4, 7) sono ingrossate più nettamente all'apice, tanto che si può tracciare un confine più o meno netto tra la parte apicale ingrossata della clava e il resto dell'antenna come flagello. La clava, a sua volta, può essere densa (Fig. 4, 8), con segmenti fusi o contigui, talvolta completamente non segmentata, talvolta lasca (Fig. 4, 9), quando i segmenti sono nettamente separati tra loro. Lamellare (Fig. 4, 10) è una mazza formata da segmenti lamellari allungati su un lato, che possono allontanarsi a ventaglio (ad esempio nello scarabeo di maggio). A volte
7 INTRODUZIONE 7Fig. 2. Vista generale del maschio dello scarabeo equino (Cicindela) dal basso. Le gambe anteriore e centrale destra sono state rimosse insieme alla coxa, quella posteriore è stata rimossa senza la coxa. (Secondo Ganglbauer con modifiche). md mandibole, baffi delle antenne, palpi, palpi mascellari, occhi, ng labbro inferiore; pb mento; gpal palpi labiali; gola monti; pgr protorace; epst I protorace episternale; epm I epimero del protorace; bacino. vp I cavità coxale anteriore; bacino I coxa anteriore; VTG I trocantere anteriore; problemi alla parte anteriore della coscia; obiettivo davanti alla tibia, zampe davanti al tarso, quando davanti agli artigli; sgr petto medio; epst II mediotorace episternale; epm II epimero del metatorace; bacino II coxa media; trocantere medio VTG II; letto II metà coscia; obiettivo II tibia media; zampa II tarso medio; ingranaggio II artigli centrali; petto posteriore zgr; epst III torace episternale; epl epipleura ed elitre; bacino III coxa posteriore; girevole posteriore VTG III; guai III coscia posteriore, obiettivo III tibia posteriore; zampe III tarso posteriore, tog III artigli posteriori; st I st VII I VII sternite addominale; penna penna è. Riso. 3. La testa dello scarabeo stafilino, Staphylinus. (Secondo Ganglbauer con modifiche). 1 vista dall'alto 2 vista dal basso md della mandibola, antenne antennali, fossa antennale; ous segmento basale dell'antenna (non conta il numero di segmenti!); palpi mascellari chshup; vg labbro superiore; g palpo palpi labiali; pb mento; cm sottomento; fascia di contanti; l fronte; t corona; guance guancia; occhio occhio; vis tempio; angolo temporale visivo; collo; gola; gola gsh; linea infraorbitaria pgl.
8 8 COLEOTTERI O SCARABEO fig. 4. Coleotteri. Tipi di struttura delle antenne. (Secondo Jacobson con modifiche). 1 filiforme (Obrium); 2 setole covid (Bembidion); 3 perline (Risodi); 4 a dente di sega (Synaptus); 5 a pettine (Rhipidius); 6 ispessimento (Bius); 7 a forma di clava (due punti); 8 clavati con mazza densa (Cruphalus); 9 clavati con mazza sciolta (Anisotoma); 10 lamellari clavati (Polyphilla); 11 lamellari a gomito (Lucanus); 12 clavato-articolato (Otiorrhynchus); 13 clavati con mazza avvolgente (Lethrus); 14 Cerocoma sbagliato); 15 irregolari, a forma di orecchio (Gyrinus). i segmenti della clava entrano l'uno nell'altro (clava avvolgente, Fig. 4, 13). Le antenne genicolate (Fig. 4, 11, 12) differiscono dai tipi elencati in quanto il 1o segmento (peduncolo) è fortemente allungato e talvolta ispessito, mentre il resto (flagello) si trova ad angolo rispetto ad esso; le antenne a gomito di solito hanno una mazza (ad esempio nei cervi, nei karapuzik, nei tonchi). Le antenne di una forma più complessa ed evasiva sono chiamate irregolari (Fig. 4, 14, 15). Seno. Dei tre segmenti toracici dei coleotteri, il protorace è il più sviluppato e il mesotorace è il meno sviluppato. Il protorace è articolato in modo mobile con il mesotorace; il mesotorace è saldato in modo inamovibile al metatorace. Ciascuno di questi segmenti è diviso nella parte superiore della schiena, nella parte inferiore del seno, o torace, e nelle parti laterali del barile, o pleura; le parti laterali del protorace sono chiamate episterne protoraciche o propleure. Il pronoto è molto variabile nella forma e nella scultura; la sua struttura è di grande importanza nella pratica di identificazione dei coleotteri. Sul pronoto si distinguono: angoli da anteriore a posteriore, che possono essere acuti, ottusi o arrotondati, bordo anteriore (apicale), bordo posteriore (bordo principale) e lati, che possono essere diritti, arrotondati, dentellati, ecc. e solitamente hanno un bordo laterale affilato. La forma del pronoto è trasversale (quando la sua larghezza è maggiore della lunghezza), longitudinale (lunghezza maggiore della larghezza), quadrata, rotonda, semicircolare, a cappuccio (Fig. 5, 1 5), ecc.; spesso ha forma di cuore, quando i suoi lati sono arrotondati nella parte anteriore, e dentellati o paralleli nella parte posteriore (Fig. 5, 6). I bordi del pronoto sono spesso bordati (delimitati), cioè separati dalla sua parte centrale (disco) da una linea depressa con un bordo. Parti dei margini laterali del pronoto ripiegati verso la parte inferiore dell'epipleura pronotale sono solitamente separate dall'episterna protoracica mediante una sutura; raramente non vi è sutura e il margine laterale del pronoto è levigato (nei tonchi, ecc. ). Il protorace ha talvolta un collare semicircolare o triangolare davanti, o una sporgenza della gola (ad esempio, in molti coleotteri Fig. 12, 5, arachidi Fig. ??, 5), che, con la testa piegata, copre l'apparato boccale da sotto. La parte posteriore del protorace talvolta sporge all'indietro tra le coxae anteriori sotto forma di un processo posteriore del protorace, che spesso (nei coleotteri clic, trivellatori, ecc.) poggia su una fossa speciale sul mesotorace. Spesso sul lato inferiore del protorace sono presenti scanalature per l'inserimento delle antenne (nei coleotteri clic, ecc.). Il grado di mobilità del protorace rispetto al mesotorace è molto diverso; a volte sono saldati quasi immobili (nei pesci rossi, nel karapuzik, ecc.), in altri il movimento è possibile solo su e giù (ad esempio nei coleotteri clic); il più mobile in tutte le direzioni è il torace anteriore di alcuni coleotteri predatori, per esempio Scaritini da coleotteri terricoli, Staphylinidae, Cleridae, ecc. Il mesotorace è ristretto anteriormente e in parte spinto nel protorace, formando il collo del mesotorace; espanso posteriormente. Porta il primo paio di ali di elitra, che sono completamente
9 INTRODUZIONE 9Fig. 5. Coleotteri. Tipi di struttura del pronoto. (Originale). 1 trasversale; 2 longitudinali; 3 quadrati; 4 rotondi; 5 incappucciati; 6 a forma di cuore. chiudilo dall'alto, ad eccezione di una piccola sezione centrale dello scudo. Dal basso è sempre più corto e spesso più stretto del metatorace; le sue episterne sono talvolta fuse con gli epimeri. Il metatorace porta il secondo paio di ali. Dall'alto è completamente ricoperto dalle elitre a riposo, e la sua parte superiore del metanoto è sempre membranosa; episterna metatoracica spesso fusa con epimeri. Sul petto di ciascun anello toracico sono presenti una coppia di cavità coxali; includono la coxa della gamba. Le cavità coxali anteriori possono essere chiuse se la coxa è circondata su tutti i lati da parti del seno e dei barili (Fig. 13, 5), e aperte quando c'è uno spazio tra le parti del seno e i barili dietro la coxa (Fig. .13, 2); le cavità coxali media e posteriore sono sempre aperte. N circa ge sempre ben sviluppato. Ciascuno di essi è costituito da cinque parti principali: coxa, trocantere, coscia, tibia e tarso. Le coxa sono poste nelle cavità coxali; la forma delle coxae è di grande importanza per l'identificazione. La coxa anteriore e quella media sono sempre mobili, sono sferiche (Fig. 13, 2), a forma di cono (Fig. 13, 1), a forma di cilindro sporgente o reclinato (Fig. 13, 4). Con le cavità coxali spalancate delle gambe anteriori, meno spesso centrali, una piccola appendice chitinosa trocantina è talvolta attaccata alla coxa. Le coxa posteriori sono solo in rari casi sferiche o sporgenti, generalmente trasversali, talvolta (ad esempio nei nuotatori) fortemente espanse; i coxa posteriori sono generalmente mobili solo in misura limitata e solo in una direzione, e talvolta (nei coleotteri, nuotatori, trivellatori, coleotteri clic, ecc.) sono immobili e saldati al metatorace. Spesso una parte della coxa posteriore si estende sotto forma di opercolo femorale sopra il femore, che può essere parzialmente o completamente nascosto sotto di esso (Fig. 11, 5); negli Haliplidae l'opercolo femorale raggiunge proporzioni enormi, ricoprendo quasi tutto l'addome. A ciascuna coxa è attaccato un piccolo trocantere, al quale di solito si unisce obliquamente, meno spesso direttamente, e il femore si muove insieme ad esso. La parte inferiore della gamba è articolata in modo mobile con la parte superiore della coscia; questa articolazione è chiamata ginocchio; la tibia è spesso ricoperta da file di spine immobili, e all'apice porta 1 2 speroni più grandi e mobili. All'apice della tibia è attaccato un tarso (raramente un po' lontano da esso), costituito nel tipo da 5 segmenti; il numero dei segmenti può essere ridotto a 4, 3, talvolta anche 2 o 1. L'ultimo segmento del tarso porta all'apice un paio di artigli, generalmente piuttosto distanziati, talvolta ravvicinati; gli artigli sono solitamente immobili, a volte possono muoversi in direzione verticale; spesso gli artigli portano un dente o un intero pettine dal basso; a volte sono divisi; in alcuni casi, un artiglio è più piccolo dell'altro o addirittura scompare completamente; a volte gli artigli si fondono alla base e anche su tutta la lunghezza; talvolta sono ridotte a semplici setole; talvolta assente del tutto. Tra gli artigli è talvolta presente un'appendice mobile empodica, solitamente a forma di lobo con 2 setole. La struttura delle zampe è di grande importanza per la definizione; i coleotteri hanno i seguenti tipi di struttura. Le gambe da corsa sono caratterizzate dall'armonia di tutte le parti e da una zampa sottile con b. M. con segmenti cilindrici (ad esempio, in molti coleotteri macinati, coleotteri scuri, ecc., Fig. 6, 1). Le gambe che camminano differiscono da loro solo per la struttura del tarso, che è espanso e appiattito, la pianta dei segmenti è densamente ricoperta di peli, il penultimo segmento è bilobato, l'artiglio è sottile, non espanso (nei taglialegna, negli scarabei fogliari, punteruoli, coccinelle, ecc.); nei tarsi di questo tipo, infatti, il 2° o 3° segmento è solitamente allargato, ed il segmento successivo (preapicale) è molto piccolo, nascosto nel precedente, e distinguibile solo sotto forte ingrandimento (falso tarso pentasegmentato). Nella gamba che scava, la gamba inferiore è espansa e il suo bordo esterno è dotato di denti (in lamellare, karapuzik, alcuni coleotteri macinati, ecc., Fig. 6, 2); il più delle volte a scavare sono le zampe anteriori. La gamba che nuota è caratterizzata da tibia e tarso estesi, i cui bordi sono ricoperti da peli fitti e forti; artigli piccoli, leggermente ricurvi
10 10 COLEOTTERI O SCARABEO fig. 6. Coleotteri. Tipi di struttura delle gambe. (Secondo Jacobson con modifiche). 1 gamba che corre (Pterostichus); 2 zampe da scavo (Scariti); 3 tappe di nuoto (Cybister); 4 zampe che saltano (argopus). tu sei e; nuotare sono le zampe centrali e posteriori degli scarabei acquatici (Fig. 6, 3). Le zampe posteriori con una coscia potente e fortemente ispessita, adattata al salto, sono chiamate salti, ad esempio, in Halticinae dai coleotteri fogliari, Rhynchaenus dai tonchi, Scirtus, ecc. (Fig. 6, 4). Il paio di ali anteriori, o elitre, hanno solitamente la stessa durezza della copertura chitinosa generale del corpo. A riposo ricoprono il mesoscuto (ad eccezione dello scutello), l'intero metanoto e la parte superiore dell'addome, ma a volte sono in una certa misura accorciati e lasciano aperti da 1 a 7 tergiti addominali. Talvolta le elitre sono fortemente sottosviluppate, e nelle femmine di alcuni Lampyrinae, Drilinae, Pachypus, Rhipidius ed altri sono completamente assenti; talvolta invece superano la lunghezza dell'addome (ad esempio nella Cassida). La linea lungo la quale le elitre si toccano è chiamata sutura e la sua parte apicale è chiamata angolo di sutura. Negli scarabei privi di ali posteriori, le elitre possono crescere insieme lungo la cucitura; a volte invece non si toccano strettamente lungo la sutura (elitre aperte), ancor più raramente (in Meloe) un'elitre si sovrappone leggermente all'altra. Sull'elitra si distinguono: il margine principale, il margine esterno o laterale (morfologicamente anteriore), l'apice (cioè l'estremità posteriore) e l'angolo omerale. Il margine principale dell'elitra è detto orlato, o bordato, se tra lo scutello e l'angolo omerale passa una linea depressa. Vicino all'angolo della spalla nei coleotteri, soprattutto quelli volanti, c'è b. M. tubercolo omerale distinto; più raramente, nella parte posteriore dell'elitra è presente anche un tubercolo preapicale (in alcuni coleotteri, punteruoli, ecc.). Il margine laterale dell'elitra è solitamente piegato sul lato inferiore, coprendo parzialmente le parti laterali del mesotorace, del metatorace e dell'addome e formando una b. M. stretta striscia di epipleura, separata dal resto dell'elitra da una curvatura distinta; in alcuni casi (in molti coleotteri oscuri), i lati più larghi delle elitre sono ripiegati sul lato ventrale, formando false epipleure, lungo il bordo delle quali sono visibili vere epipleure più strette. La scultura delle elitre è molto importante per l'identificazione dei coleotteri; allo stesso tempo, la venatura è visibile sulla superficie superiore solo in alcuni coleotteri (Lycinae, Arrhaphipterus), ma di solito dall'alto è visibile un'ampia varietà di strutture, da una superficie liscia e lucida a varie chiglie, spine, escrescenze e fosse. Molto spesso sulle elitre sono presenti solchi o file longitudinali b. M. punti profondi, o solchi punteggiati, separati l'uno dall'altro da spazi vuoti. Questi spazi possono essere sollevati sotto forma di chiglie o costole, e le chiglie o le costole possono essere spezzate in file di tubercoli o grani, o interrotte da file di fosse. Si contano i solchi e gli spazi per ciascuna elitra dalla sutura al margine esterno; il primo solco è chiamato sutura. Vicino allo scutello è spesso presente un altro breve solco, lo scutello, che non è compreso nel conteggio (Fig. ??, 17). Gli intervalli dispari (1, 3, 5, 6, 9) a volte differiscono nella scultura da quelli pari (2, 4, 6, 8). Gli apici di entrambe le elitre piegati insieme formano molto spesso un arco comune, ma a volte l'angolo di sutura sporge a forma di dente o spina dorsale o è allungato a forma di coda, oppure ciascuna elitra è arrotondata ma separatamente; spesso l'apice delle elitre è troncato o tagliato rettilineamente, in modo che dietro di esso sia visibile l'apice esposto dell'addome, e talvolta una parte più ampia di esso (elitre accorciate). Le ali posteriori (inferiori) (di seguito semplicemente ali) vengono utilizzate per il volo; sono palmati, b. M. trasparente, con venature più scure. Le ali possono essere ridotte o del tutto assenti, negli stessi casi in cui sono adatte al volo, sono un po' (a volte notevolmente) più lunghe e larghe delle elitre, ma a riposo completamente nascoste sotto di esse (fanno eccezione Necydalis, Molorchus, Rhipiphorus e pochi altri), non importa quanto corte siano le elitre (ad esempio negli stafilinidi); mentre le ali sono piegate in senso trasversale e pro-
11 INTRODUZIONE 11 Fig. 7. Coleotteri. Dettagli costruttivi. (Secondo Jeannel, Cult, Lindroth e Jacobson). 1 venatura di tipo caraboidale (Asaphidion); 2 venature di tipo stafilinoide (Nicrophorus); 3 tipo di venatura cantaroide (Lygistopterus); 4 ali pennate (Ptilium); 5 pori contenenti setole (notevolmente ingranditi); 6 microsculture isodiametriche; 7 microsculture a maglie incrociate; 8 microsculture incrociate striate. direzione longitudinale. Per il volo gli scarafaggi sono soliti sollevare le elitre ed estendere le ali, allargandole; pochi coleotteri (coleotteri bronzei, Gymnopleurus) possono volare senza sollevare le elitre per la presenza di apposite tacche sui lati di queste ultime. La disposizione delle vene sulle venature delle ali gioca un ruolo importante nella moderna classificazione dei coleotteri. Esistono tre tipi principali di venatura alare nei coleotteri: 1) tipo caraboid (Fig. 7, 1); caratterizzato dalla presenza di 1 2 vene trasversali tra i rami della vena mediale (M 1 e M 2), che formano una cellula oblunga chiusa; 2) tipo stafilinoide (Fig. 7, 2); si distingue per la completa assenza di cellule trasversali e per il sottosviluppo della base del ramo M 1, che non è collegato al tronco principale M; 3) tipo cantaroide (Fig. 7, 5), il più comune; caratterizzato dal fatto che la parte principale della vena M 2 forma una vena ricorrente, che si collega allo stelo principale con un'inflessione a forma di uncino (a volte molto breve, e in alcuni casi scompare completamente, in modo che la venatura assomiglia quindi al tipo stafilinoide). I tipi di venatura descritti sono caratteristici di interi sottordini e superfamiglie di coleotteri (vedi Chiave delle famiglie). In alcuni gruppi dei coleotteri più piccoli (Ptiliidae, Sphaeriidae, ecc.), La placca alare è molto poco sviluppata, ma lungo i bordi è presente una frangia di peli lunghi (Fig. 7, 4), con l'aiuto del quale volo impennato viene effettuata. L'addome è strettamente adiacente al metatorace e alla coxa posteriore. Il numero totale di segmenti addominali non supera 10 e ciascuno di essi è costituito da due semianelli: dorsale (tergite) e addominale (sternite). Il numero di sterniti visibili è solitamente 5 7, e il numero di tergiti 7 9, poiché gli sterniti dei primi 2 segmenti sono solitamente assenti nei coleotteri, mentre gli ultimi 1 3 segmenti vengono modificati e, di regola, vengono spinti all'interno addome. I tergiti ricoperti dalle elitre sono debolmente chitinizzati, mentre quelli non ricoperti da esse sono duri come gli sterniti (ad esempio nelle stafiline). Gli spiracoli ventrali si trovano sulle membrane tra i tergiti e gli sterniti. L'ultimo (apicale) dei tergiti esposti è detto pygidium, ed il penultimo è detto propygidium; in molti casi il pygidium è situato verticalmente (in molti arachidi, lamellare, cariossidi, in alcuni coleotteri fogliari). In molti gruppi di coleotteri lo sternite I è ricoperto al centro dal metatorace e dalla coxa posteriore, sicché di esso rimangono solo le parti laterali. A volte gli sterniti hanno solchi ventrali trasversali depressi. L'ultimo sternite (apicale) è chiamato anale; la sua struttura è spesso diversa nei diversi sessi. Il colore degli scarafaggi è molto vario; può dipendere dalla presenza di pigmenti nel tegumento del corpo (toni non metallici) o da una particolare struttura microscopica (tonalità metalliche), o, infine, da una combinazione di pigmenti e strutture. I colori strutturali e alcuni pigmenti (soprattutto il marrone e il nero) sono molto resistenti, mentre i toni non metallici del rosso e del verde sono generalmente meno stabili e dopo la morte dello scarabeo cambiano colore in giallo, giallo-marrone o rosso; questi cambiamenti di colore post mortem devono essere presi in considerazione nell'identificazione (specialmente in Cassida, Mylabris e alcuni altri).
12 12 COLEOPTERI O SCARABEO La colorazione può essere monocolore o consistere in una combinazione di diversi, molto spesso 2 3, raramente più colori. Il colore della parte superiore del corpo è particolarmente vario. Sullo sfondo generale di un colore potrebbero esserci macchie o un altro motivo di un altro colore. Se le macchie sono allungate sotto forma di strisce, si distinguono strisce longitudinali e bande trasversali. Le macchie possono anche essere oculari a forma di anello. Nella stessa specie, la colorazione può variare notevolmente: in alcuni casi, lo scarabeo (o qualsiasi parte di esso) può essere monocolore con macchie chiare o scure o addirittura completamente scure, e i toni metallici possono essere rappresentati da tutte le sfumature da dal nero-blu al rosso rame. Macchie e strisce possono scomparire o espandersi e fondersi tra loro. Nonostante tale incostanza, ogni specie ha un certo tipo di colorazione, dalla quale può essere distinta dalla posizione delle macchie e delle strisce e dal modo in cui cambiano. La superficie del corpo può essere liscia, talvolta quasi lucida e irregolare o ruvida; soprattutto spesso è coperto di punti o rughe, le cui dimensioni e densità sono molto diverse, inoltre possono essere combinate. Sulla superficie possono essere presenti anche vari rilievi di granuli, tubercoli, spine, ecc. In alcuni casi è importante per la determinazione la microscultura, visibile sotto ingrandimento (in alcuni casi) su superfici che appaiono lisce a un ingrandimento inferiore. Esistono tre tipi principali di microscultura: isodiametrica (Fig. 7, 6), reticolata incrociata (Fig. 7, 7) e striata trasversalmente (Fig. 7, 8); quest'ultimo tipo di microscultura provoca spesso una lucentezza iridescente della superficie. Oltre alle tipologie di irregolarità elencate, che sono semplici depressioni o rilievi della superficie, esistono formazioni ancora più complesse articolate con la superficie: squame, peli, setole. La loro densità può essere molto diversa; le squame sono dipinte in una varietà di colori; i peli variano dal bianco e grigio chiaro passando per il giallo, grigio e bruno fino al nero; raramente i peli sono metallici. La differenza tra peli e setole è che i peli sono chiamati sottili, b. M. formazioni morbide, non ristrette alla base e solitamente sedute in una piccola depressione (punto) o su un piccolo tubercolo; setae gli stessi peli, ma più spessi e più duri, sporgenti b. M. verticalmente e attaccato in uno speciale approfondimento del poro portasetole (Fig. 7, 5). La disposizione delle sete (chetotassia) è molto costante e gioca quindi un ruolo importante nell'identificazione (soprattutto di coleotteri e stafilinidi); se le setole si sono rotte o sono cadute, il punto in cui sono attaccate può essere determinato dalla presenza di un poro portasetole. Oltre a queste strutture, il corpo di molti coleotteri durante la vita è ricoperto di polline o placca, che di solito vengono cancellati molto facilmente, ma possono essere ripristinati negli individui viventi. I peli, le scaglie e il polline possono essere così densi da nascondere completamente il colore sottostante; spesso formano anche macchie o strisce. Differenze sessuali esterne (dimorfismo sessuale). Oltre alla struttura degli organi genitali (vedi sotto), i diversi sessi della stessa specie di coleottero spesso differiscono notevolmente nell'aspetto. I maschi sono generalmente un po' più piccoli e hanno un corpo più stretto rispetto alle femmine; talvolta i maschi sono più grandi (con ornamenti molto sviluppati sulla testa e sul pronoto, ad esempio nei Lucanidae, in alcuni Dynastinae, Lethrus, Labidostomis), e occasionalmente i maschi hanno un corpo più largo (in alcuni Endomychidae). Spesso l'addome del maschio ha un'impronta longitudinale, mentre la femmina ha un'impronta convessa (ad esempio in Cetonia aurata L.). Molto spesso (nella maggior parte dei coleotteri e dei nuotatori, in molti coleotteri scuri, coleotteri fogliari, alcuni coleotteri, ecc.), le zampe anteriori e talvolta quelle centrali del maschio sono molto espanse e portano una suola di peli aspiranti, che serve a tenere la femmina durante l'accoppiamento; oltre a questo, le femmine di molti coleotteri acquatici hanno una speciale scultura solcata e ruvida, che le rende più facili da impugnare da parte dei maschi. Spesso i maschi hanno femori posteriori gonfi e talvolta spinosi o tibie anteriori, medie o posteriori ricurve e spesso allungate; a volte ci sono punte o tacche sugli stinchi dei maschi. Spesso i maschi hanno mandibole molto sviluppate (nei Lucanidae, Lethrus, Labidostomis, ecc.). In molti casi, la testa e il pronoto del maschio portano varie corna, escrescenze, tubercoli, ecc. Molto spesso la lunghezza delle antenne o lo sviluppo dei processi sui segmenti, o la dimensione della mazza, è maggiore nei maschi che nelle femmine; in alcuni casi, i maschi hanno ancora più segmenti sulle antenne rispetto alle femmine. A volte i maschi hanno palpi notevolmente ingranditi; pare che in tutti questi casi si ottenga un aumento della superficie degli organi olfattivi, che facilita la ricerca delle femmine. Gli occhi allargati del maschio (ad esempio nelle Lampyrinae) servono a facilitare similmente la ricerca delle femmine.
13 INTRODUZIONE 13 Fig. 8. Coleotteri. (Secondo Lindroth e org.). 1 organi genitali del maschio Elater (sclera basale, sclerite basale, pene del pene, parameri); 2 dissezione dei genitali. Lo stile di vita meno mobile delle femmine porta ad una più frequente assenza delle ali inferiori; tali sono le femmine senza ali di Ptino. molti coleotteri, alcuni boscaioli, ecc., mentre i maschi di queste specie sono alati. Talvolta le elitre delle femmine sono sottosviluppate o addirittura del tutto assenti (nelle lucciole, Drilus, Rhipidius); in casi molto rari (in alcuni scolitidi), invece, le femmine sono alate, mentre i maschi hanno ali ridotte. In alcuni casi, i pavimenti differiscono per colore o struttura della superficie. Esistono numerosi segni di dimorfismo sessuale, che vengono specificati, se necessario, nelle tabelle chiave. In molti casi, per determinare le specie di coleotteri, è necessario studiare le parti chitinose degli organi genitali (genitali) del maschio. Questi organi sono tipicamente costituiti dall'organo copulatore vero e proprio (pene), da 2 parameri con esso articolati e da uno sclerite basale (Fig. 8, 1). Spesso (negli Scarabaeidae, Histeridae, Tenebrionidae, Chrisomelidae, e molte altre famiglie) i parameri si fondono tra loro, formando un tubo più o meno chiuso, all'interno del quale si trova il pene. Per studiare i genitali è necessario sollevare le elitre, tagliare la membrana tra i tergiti addominali (Fig. 8, 2) ed estrarre i genitali con un ago da dissezione. I campioni asciutti devono essere immersi in una camera umida per 1-2 giorni prima della preparazione, oppure immersi in acqua tiepida (ma non bollente) per un breve periodo fino a quando le giunture diventano flessibili; quest'ultimo non deve essere eseguito con specie ricoperte di scaglie o peli, i genitali estratti devono essere bolliti per diversi minuti in una soluzione di idrossido di potassio (KOH) al 10% per rimuovere i tessuti molli residui, e quindi risciacquati abbondantemente con acqua pulita. I genitali sezionati vengono incorporati in una preparazione permanente o temporanea, oppure vengono incollati su un rettangolo di cartone fissato con spilli in modo da sembrare uno scarabeo. Le uova degli scarafaggi sono generalmente ovali, traslucide, di colore chiaro (il più delle volte bianche, a volte giallo chiaro, grigio o verde chiaro), il loro guscio è sottile, senza una scultura affilata. L e h i n k e con una testa ben sviluppata, fortemente sclerotizzata e apparato boccale rosicchiante. Il corpo è generalmente molle, bianco nelle forme latenti, più scuro, grigio, bruno, nero nelle forme a vita aperta; nelle larve che si nutrono di foglie, talvolta verdi; a volte ci sono macchie gialle o rosse brillanti sullo sfondo principale. In alcuni gruppi, il corpo della larva, soprattutto dall'alto, è fortemente sclerotizzato (nei coleotteri clic, nei coleotteri scuri, in alcuni coleotteri macinati) e è colorato di giallo, marrone o nero, molto raramente blu metallizzato o verde sopra. Secondo la struttura, si possono distinguere diversi tipi principali di larve di coleottero. Le larve campodeiformi (Fig. 9, 1) hanno 3 paia di lunghe zampe toraciche, antenne distinte e spesso filamenti caudali, i loro corpi sono allungati e molto mobili (ad esempio nei coleotteri, negli stafilinidi, nei nuotatori). Le larve erucoidi (Fig. 9, 2) sono simili a vermi, con zampe corte o senza zampe, con antenne corte, appena percettibili, dalla pelle morbida e inattive (ad esempio, nei lamellari, nei taglialegna, nei tonchi). Le larve del tipo "wireworm" (Fig. 9, 3) hanno un corpo giallo o marrone lungo, cilindrico, completamente fortemente sclerotizzato con zampe corte e forti (nei coleotteri clic, coleotteri scuri, coleotteri pollinici). Esistono anche altri tipi di larve: fortemente pelose (nei coleotteri cutanei), piatte (in Silpha e altri), ricoperte da lunghe escrescenze respiratorie (branchie tracheali delle larve di Haliplidae che vivono nell'acqua). In alcuni casi, c'è ipermetamor-
14 14 COLEOTTERI O SCARABEO fig. 9. Coleotteri. Principali tipi di larve. (Secondo Jacobson). 1 campodeiforme (Carabidi); 2 erucoidi (scarabaeidi); 3 "verme metallico" (Elateridae). phosis (vedi t. I), ad esempio, nei coleotteri vescicolari, campodeiformi, larve molto mobili del 1° stadio penetrano nelle capsule degli acridoidi o nei nidi degli imenotteri e successivamente si trasformano in larve erucoidi e inattive del 2° stadio. L'impupamento avviene più spesso nel terreno, sotto la corteccia o nel legno degli alberi in cui vivevano le larve. Per la pupa le larve preparano la cosiddetta "culla". La maggior parte delle larve acquatiche si impupano anche nel terreno (lungo le sponde). Poche larve tessono un vero e proprio bozzolo (Phytonomus, Cionus). Le larve delle coccinelle e di alcuni coleotteri fogliari si impupano apertamente sulle piante, attaccandosi ad esse con l'estremità posteriore del corpo. Le pupe che si sviluppano nelle culle sono libere, morbide, solitamente bianche, ed è facile riconoscere in esse tutte le parti del futuro coleottero. Le pupe apertamente attaccate sono spesso dai colori vivaci e le loro coperture sono più fortemente sclerotizzate. La biologia degli hardwing è estremamente diversificata; sotto questo aspetto superano quasi tutti gli altri ordini di insetti. Tra questi ci sono rappresentanti della maggior parte dei principali tipi di alimenti conosciuti nella classe degli insetti. I predatori sono numerosi, sia polifagi che specializzati. Esistono forme ancora più erbivore (fitofagi), tra cui i consumatori di fogliame (fitofagi propriamente detti), radici (rizofagi), legno e corteccia (xilofagi), fiori (antofagi), frutti e semi (carpofagi). La maggior parte dei fitofagi si sviluppa su tessuti viventi, ma alcuni sono in grado di vivere nel legno secco. Vasti gruppi biologici sono consumatori di funghi (micetofagi); specie che si nutrono di animali in decomposizione e di materia vegetale (necrofagi e saprofagi); detritifagi che si nutrono di materia vegetale e animale secca o in lenta decomposizione. Questi gruppi sono collegati da molte transizioni. I coleotteri abitano le terre e le acque dolci dell'intero globo, dai deserti più aridi alle foreste tropicali, dalle regioni equatoriali alla tundra. Sono assenti solo nelle zone ricoperte da una calotta glaciale permanente (Antartide, alcune zone dell'Artico e le parti più alte delle catene montuose). Nel Caucaso, gli scarabei raggiungono un'altezza di oltre 3000 m, nelle montagne dell'Asia centrale fino a 4500 m, e nell'Himalaya e nelle montagne dei tropici fino a M. Nelle regioni abitate dagli scarafaggi, sono trovato quasi ovunque. Moltissimi di loro vivono negli strati superiori del suolo, e le larve e le pupe trascorrono lì tutta la loro vita, e gli adulti nella maggior parte dei casi vengono in superficie almeno per un breve periodo (ad eccezione di alcuni piccoli coleotteri terricoli, rove coleotteri, punteruoli, ecc.). Non meno numerosi sono gli abitanti delle piante. Alcuni vivono sotto la corteccia e nel legno, altri sulle foglie (e talvolta all'interno dei loro tessuti), nei fiori e nei frutti, altri ancora nelle radici e su di essi. Le piante vengono cacciate anche da molti coleotteri predatori (coccinelle, alcuni coleotteri terricoli). I rappresentanti di numerose famiglie vivono di alberi e funghi terrestri, nutrendosi di essi (micetofagi) o cacciando altri animali; tali forme di funghi sono particolarmente caratteristiche delle aree forestali umide. Moltissimi coleotteri vivono nel letame o nelle carogne, alcuni qui si nutrono di sostanze in decomposizione, altri predano, in particolare a spese delle larve di mosca. Nei corpi d'acqua dolce vive una fauna molto ricca di coleotteri; alcuni di loro appartengono a famiglie specificamente acquatiche (galleggiante, vorticoso, idrofilo, Driopidi, ecc.), altri sono rappresentanti specializzati di famiglie che vivono principalmente sulla terra (ad esempio, i coleotteri delle foglie dell'iride della sottofamiglia Donaciinae, alcuni tonchi). Pochi
16 16 COLEOPTERI O SCARABEO sia il lungo sviluppo (diversi anni) della larva che la lunga vita dell'adulto. La fase ibernata è solitamente quella di uno scarafaggio o di una larva, mentre le uova e le pupe vanno in letargo in rare occasioni. L'importanza degli scarafaggi per l'uomo è determinata principalmente dal fatto che tra loro esiste un numero significativo di parassiti primari dell'agricoltura e della silvicoltura. Così, tra i parassiti delle colture in campo dovrebbero essere nominati lo scarabeo della patata del Colorado, i punteruoli della barbabietola e alcune altre specie di questa famiglia, molte specie di coleotteri a clic, coleotteri scuri, baffi lamellari, pulci della terra del pane e del lino, scarafaggi del pane, scarabei del grano e un lungo numero di altri. Anche le colture orticole soffrono molto a causa dei coleotteri dannosi. Chiamiamo, ad esempio, le pulci di terra delle crocifere e altri tipi di coleotteri fogliari, molti tonchi, coccinella di melone. Ancora più importanti sono i coleotteri come parassiti del giardino, tra cui ricordiamo gli elefanti Attelabidae (oca, grande elefante pavone, bukarka), trivellatori (in primis il piralide nero Capnodis tenebrioides F.), alcuni taglialegna, ecc. scolitidi, nonché larve di coleotteri, che danneggiano gravemente vivai e vigneti. Il ruolo dei coleotteri come parassiti delle foreste è eccezionalmente grande. Qui gli scarabei corteccia e i boscaioli sono più conosciuti, nel sud sono conosciuti anche gli scarabei dorati. Le giovani piantagioni forestali sono gravemente danneggiate dalle larve dei coleotteri clic (wireworms) e dei coleotteri, e nelle aree di rimboschimento della steppa e in Asia centrale anche dai coleotteri scuri. È inoltre necessario tenere conto dell'attività degli scarabei come parassiti dei pascoli naturali. Notiamo, ad esempio, le riproduzioni di massa dello scarabeo fogliare dell'assenzio (Theone silphoides Dalm.), che portano a un catastrofico calo della produttività dei pascoli in Kazakistan. I pascoli ai piedi e sulle montagne dell'Asia centrale sono gravemente danneggiati da vari tipi di coleotteri oscuri, e in particolare dai kravchiki (Lethrus). Il valore negativo di molti coleotteri (finti, coleotteri cutanei, alcuni coleotteri scuri, punteruoli, scarafaggi della farina della famiglia Cucujidae, ecc.) come parassiti di ceppi di origine vegetale e animale è elevato. La maggior parte di questi parassiti sono polifagi, alcuni si nutrono solo di determinati tipi di ceppi (ad esempio, punteruolo dei fagioli e altri legumi). Molti di loro sono gli oggetti più importanti della quarantena esterna (ad esempio, lo scarabeo kapr Trogoderma granarium Ev.) o interna. Il ruolo di alcuni coleotteri, in particolare scarabei stercorari e scarafaggi scuri, merita particolare attenzione come ospiti intermedi di malattie da elminti nell'uomo e negli animali domestici, nonché come portatori meccanici di malattie gastrointestinali e invasioni da elminti. Molti coleotteri, soprattutto insetti (Meloidae), ma anche alcuni coleotteri fogliari (ad esempio Theone), coccinelle, stalinidi (Paederus), coleotteri dalle ali strette (Nacerda) contengono sostanze protettive altamente tossiche (cantaridina, ecc.) sangue dei genitali. Queste sostanze, a contatto con la pelle e le mucose, possono provocare gravi infiammazioni simili a ustioni. Il consumo di questi coleotteri da parte del bestiame (insieme alla vegetazione) può causare avvelenamento e morte degli animali. Infine, alcune specie di grandi nuotatori (Cybister, Dytiscus) possono danneggiare gravemente la piscicoltura e la pesca in generale, sterminando il novellame dei pesci commerciali. Anche il ruolo utile degli scarabei è molto grande. Numerosi predatori che sterminano gli insetti dannosi (entomofagi) dovrebbero essere nominati in primo luogo tra gli scarafaggi utili. Particolarmente conosciute sono le coccinelle (Coccinellidae), molte delle cui specie sono i più importanti nemici naturali di parassiti pericolosi come afidi, cocciniglie e acari. All'uso delle coccinelle (Rodolia cardinalis Muls., Cryptolaemus montrouzieri Muls.) sono associati numerosi seri successi nel metodo biologico di lotta contro gli insetti dannosi. Non meno importanti sono i coleotteri terricoli, predatori molto attivi, per lo più polifagi. Le più famose tra loro sono le bellezze (Calosoma), che sterminano i bruchi dei parassiti nell'agricoltura e nella silvicoltura. Negli ultimi decenni, i coleotteri terricoli hanno attirato l'attenzione in molti paesi come il principale fattore naturale che frena l'attività dannosa dei vermi e di altri parassiti del suolo e come nemici naturali dello scarabeo della patata del Colorado. Il ruolo utile di questa famiglia richiede ulteriori studi. Un ruolo importante come entomofagi è svolto da tante piccole noccioline (Histeridae). Si nutrono principalmente delle larve di altri insetti; le specie di sterco e carogne mangiano larve di mosca; quelli che vivono sotto la corteccia distruggono uova e larve di scolitidi, boscaioli, ecc.; gli abitanti delle tane e dei nidi dei roditori sterminano le larve dei ditteri (comprese le zanzare) e le pulci. Alcuni nuotatori non hanno poca importanza, sterminando le larve delle zanzare succhiasangue. Tra gli entomofagi vanno nominati
17 INTRODUZIONE 17 anche molti Staphylinidae, Cleridae, alcuni Cantharidae e anche larve di singole specie di Elateridae. Le suddette larve di molti insetti (Mylabris, Epicauta) riducono il numero delle locuste dannose sviluppandosi nei loro baccelli-uova. Un posto importante in natura come inservienti, che distruggono e seppelliscono sostanze in decomposizione (sterco, carogne) nel terreno, è occupato dagli scarabei stercorari (in particolare rappresentanti delle sottofamiglie Coprinae e Geotrupinae), scarafaggi morti e altri scarabei stercorari e carogne. Molti coleotteri, nutrendosi di fiori, svolgono un ruolo importante nell'impollinazione incrociata di piante entomofile, e alcune forme altamente pubescenti, come Amphicoma, sono apparentemente i principali impollinatori specifici di piante da fiore come i tulipani, makiidr. Infine, è necessario sottolineare l'importanza dei coleotteri come fattore più importante nella circolazione delle sostanze in natura, principalmente nella formazione del suolo e in altri processi biogeochimici. In particolare i coleotteri, che si nutrono di diverse sostanze in decomposizione, svolgono un ruolo molto importante nella mineralizzazione dei residui animali e vegetali e nell'arricchimento con essi degli strati profondi del terreno. Letteratura essenziale. F e in URSS. Coleotteri. Ed. Accademia delle Scienze dell'URSS (pubblicato in numeri separati, nel corso degli anni sono stati pubblicati 17 volumi). Sto cantando. G. Coleotteri della Russia e Zap. Europa. Pgr., ed. Devrien, 1024 pagine, 83 colori. tavole (non completate, termina la famiglia Antichidae); Sto cantando. D. Identificazione del coleottero. M. L., GIZ, 1927, 522 pagine; 2a edizione, 1931, 472 pagine; B e s t i m u n g s-ta b e l l e n d e r e ur o p a i s c h e n C o l e o p t e r e n, herausg. von E. Reitter, H, (pubblicato in numeri separati; la maggior parte dei numeri include tutti i rappresentanti paleartici di questo gruppo); G a n g l b a u e r L. Die Käfer von Mitteleuropas. bd. Io IV. Vienna, 2769 pp.; J u n k W. et S. S c h e n - k l i n g. Catalogo del Coleopterorum. Parte 1 160 (il catalogo più completo della fauna mondiale dei coleotteri; dal 1949 sono stati pubblicati numeri separati come seconda edizione); K u h n t P, Illustrierte Bestimmungs-Tabellen der Käfer Deutschlands. Stoccarda, 1912, 1127 pp., disegno; Reit t e r E. Fauna Germanica. Die Käfer des Deutschen Reichs. I V. Stoccarda, 1655 pp., 168 col. tavoli; W i n k l e r A. Catalogus Coleopterorum regionis palaearcticae. vita. 1 13, Vienna, 1698 pag.
Nome degli scarafaggi nelle immagini atlas determinant >>> Nome degli scarafaggi nelle immagini atlas determinant Nome degli scarafaggi nelle immagini atlas determinant I più grandi coleotteri del mondo Una volta durante l'infanzia ci sembrava
Atti del Centro scientifico di Chelyabinsk, n. 3(29), 2005 [e-mail protetta] Istituto di Sistematica ed Ecologia degli Animali
M.V. Chertoprud, E.S. Chertoprud UNA BREVE CHIAVE PER GLI INVERTEBRATI DELL'ACQUA DOLCE NEL CENTRO DELLA RUSSIA EUROPEA Classe Insetti Insecta
Le popolazioni di coleotteri longhorn ege vivono a lungo nelle foreste di conifere >>> Le popolazioni di coleotteri longhorn vivono a lungo nelle foreste di conifere Nelle foreste di conifere le popolazioni di coleotteri longhorn vivono a lungo
1 2. Ricorda come respirano i molluschi terrestri. Cosa hanno in comune i molluschi terrestri e i ragni? 1 3. Qual è il nome della riproduzione, in cui solo dalle uova non fecondate deposte dalla femmina
Zemoglyadchuk A.V. Adattamento morfologico delle larve degli scarabei megatteri (Coleoptera: Mordellidae) alla vita in un ambiente denso Le larve degli scarabei megatteri si sviluppano nei tessuti
Farfalla Nelle distese di prati d'erbe, nella steppa e nei campi durante il periodo di fioritura delle varie piante, si possono vedere sempre moltissime farfalle. Accarezzano lo sguardo dell'uomo, essendo un ornamento della natura. Come nel colore
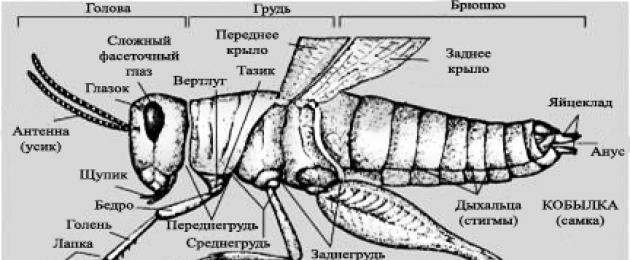
ENTOMOLOGIA SCIENZA DEGLI INSETTI ("entomos" - insetti, "logos" - scienza) E scopriamolo insieme ai ragazzi "- Chi sono questi insetti?" Tutti gli insetti hanno sei zampe La struttura esterna degli insetti. Corpo
Lucrarea de laborator 3 (523) ANATOMIA, SISTEMATICA ŞI FIZIOLOGIA ANIMALELOR (50 punti) I. ANATOMIA ANIMALE. (15 punti) 1. Esaminare i preparati proposti utilizzando un microscopio 2. Spiegare il quadro istologico
Ministero dell'Istruzione della Repubblica di Bielorussia Istituzione educativa "Università statale di Gomel intitolata a Francisk Skorina" N.G. Galinovsky, V.N. Veremeev ENTOMOLOGIA Guida pratica
Lavoro di laboratorio 1 Studio della struttura e del movimento degli animali unicellulari. Scopo: studiare le caratteristiche della struttura e del movimento degli animali unicellulari. Attrezzatura: microscopio, vetrini e coprioggetto
REVISIONE ENTOMOLOGICA, LXXIV, 4, 1995 UDC 595.792 075.1) 1995 3. A. Efremova e M. V. Kriskovich NUOVO GENERE E NUOVA SPECIE DI EULOPHIDS (HYMENOPTERA, EULOPHIDAE) DALL'UZBEKISTAN)
- In contatto con 0
- Google Plus 0
- OK 0
- Facebook 0






