Le differenze sono emisferi cerebrali del prosencefalo, che sono di dimensioni maggiori e hanno una struttura più complessa. Lo strato esterno del cervello è costituito da uno strato separato di cellule nervose che si forma corteccia cerebrale. Nella maggior parte dei mammiferi, la corteccia cerebrale è costituita da un numero così elevato di neuroni che non si trova in uno strato continuo, ma forma circonvoluzioni. Convoluzioni- Questo è il segno principale di un cervello ben sviluppato. La corteccia cerebrale è responsabile di molte funzioni, tra cui formazione di riflessi condizionati, che non si formano quando viene rimossa la corteccia.
Come il cervello anteriore, cervelletto nei mammiferiè sviluppato molto bene e ha anche delle convoluzioni. Questo livello di sviluppo cerebellare è dovuto al fatto che i mammiferi devono essere in grado di coordinare movimenti complessi e vari. Anche i mammiferi sono ben sviluppati organi di senso.
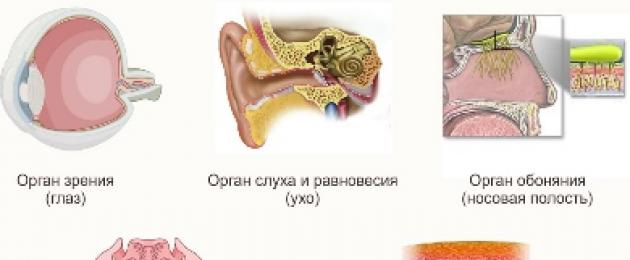
I mammiferi sono ben sviluppati organi di senso: vista, olfatto, udito, gusto e olfatto. Ma il livello di sviluppo o coinvolgimento di ciascuno di essi in una particolare specie dipende dall'habitat di questa specie di mammifero.

Ad esempio, una talpa che vive sottoterra ha una vista poco sviluppata. Le balene e i delfini che vivono nell'acqua non hanno quasi alcuna capacità di annusare, sebbene molti mammiferi terrestri abbiano un senso dell'olfatto molto acuto. Olfatto acuto per mammiferi terrestri- questo è il principale organo di senso che ti permette di trovare la preda, percepire l'avvicinarsi di un predatore e trovare una femmina o un maschio per l'accoppiamento. I genitori identificano i loro bambini dall'olfatto.
Anche l’udito gioca un ruolo importante per la maggior parte dei mammiferi. Per rilevare i suoni meno evidenti, i mammiferi hanno orecchie, mobile nella maggior parte di essi. Inoltre, in molti animali per i quali l'udito è molto importante, la superficie del padiglione auricolare è spesso ricoperta di peli sensibili alle più piccole vibrazioni sonore - vibrisse.
La vista dei mammiferi non è così acuta come quella degli uccelli; molti mammiferi sono parzialmente o completamente incapaci di distinguere i colori. Solo le scimmie possono distinguere la stessa gamma di colori degli esseri umani.
Gli organi del tatto sono vibrisse, che crescono su parti sensibili del corpo, come la testa. Anche le scimmie, come gli esseri umani, usano la punta delle dita per il tatto. Boccioli di gusto ben sviluppato negli erbivori, grazie al quale distinguono facilmente le piante commestibili da quelle non commestibili. A causa della complessità del sistema nervoso comportamento dei mammiferi non è meno complesso di quello degli uccelli e di altri animali.
Comportamento dei mammiferi.
Il comportamento dei mammiferi è complesso a causa di diversi fattori:
Prosencefalo e corteccia cerebrale ben sviluppati, nonché sistema nervoso generalmente;
La capacità di produrne molti riflessi condizionati per tutta la vita.
Fin dai primi giorni di vita, i bambini sono in grado di identificare la madre. Man mano che crescono, aumenta l'esperienza delle relazioni tra gli individui e l'ambiente esterno, acquisiscono conoscenza nel processo Giochi: saltare, combattere, cacciare, inseguire, correre, ecc.). Questi giochi aiutano anche a svilupparsi competenze, migliorando la resistenza dei bambini. In futuro, queste abilità aiuteranno anche i mammiferi a cacciare o a salvarsi la vita.
Nel corso della loro vita, i mammiferi sviluppano un gran numero di riflessi condizionati dovuti alla variabilità ambientale. Vecchi riflessi condizionati che non sono rinforzati stimoli condizionati, possono andare perduti nel tempo, il che ne consente un utilizzo razionale risorse cerebrali.
§ 49. L'emergere del cervello dei mammiferi
Piccoli antenati rettiliani di mammiferi emersero dai detriti legnosi del Carbonifero con un senso dell'olfatto sviluppato, apparato vestibolare, vista scarsa e centri di associazione nel mesencefalo. Queste creature iniziarono un misterioso percorso evolutivo che da quasi 60 milioni di anni non è stato segnato da chiare tracce paleontologiche. I triconodonti apparvero solo nel tardo Triassico (Megazostrodonte), che possono essere considerati mammiferi antichi, ma completamente sviluppati. Nel corso di diverse decine di milioni di anni si verificarono eventi che portarono alla formazione di un perfetto sistema associativo del prosencefalo, del sangue caldo, dello sviluppo placentare e dell’alimentazione dei piccoli con il latte (Kemp, 1982; Tyndale-Biscoe, Rentree, 1987).
Proviamo a valutare i cambiamenti nel sistema nervoso che hanno preceduto la comparsa dei triconodonti. Gli antenati dei mammiferi del Carbonifero possedevano una serie di qualità caratteristiche della maggior parte dei rettili di quel periodo. Per diventare mammiferi dovevano trovarsi in un ambiente in cui le loro caratteristiche morfofunzionali offrissero i massimi vantaggi biologici.
La maggior parte dei mammiferi moderni ha un senso dell'olfatto sviluppato. Si perde secondariamente negli odontoceti e viene utilizzato relativamente poco dai proboscidati, dai pipistrelli e dai primati. In altri casi, i mammiferi utilizzano ampiamente sia l'organo principale dell'olfatto che il sistema vomeronasale. Nei mammiferi più primitivi l'olfatto gioca un ruolo di primo piano e la rappresentazione dei centri chemocettori nel prosencefalo può superare tutte le altre strutture messe insieme (vedi Fig. III-19, a). È ovvio che nelle prime fasi dell'evoluzione dei mammiferi l'olfatto giocava un ruolo importante. Questa era la ragione dello sviluppo preferenziale degli emisferi del prosencefalo. Lo sviluppo del sistema olfattivo ha dato origine agli emisferi del prosencefalo, che dominano il resto del cervello. Il volume degli emisferi accoppiati dei mammiferi è sempre maggiore del volume di altre strutture del sistema nervoso, indipendentemente dalla specializzazione di una particolare specie (vedi Fig. III-18; III-19; III-21; III-25) .
Lo sviluppo dell'olfatto e del proencefalo è stato il primo grande evento neurologico nella storia di questo gruppo. Si può presumere che gli antenati dei mammiferi utilizzassero l'olfatto come principale sistema di afferentazione. A quali condizioni ciò potrebbe accadere? La situazione ovvia è l'attività notturna dei mammiferi arcaici, ma l'udito, la vista, il tatto e i termorecettori possono essere utilizzati con successo per la caccia notturna. I mammiferi preferivano usare l'olfatto, sebbene gli altri sensi non fossero significativamente ridotti.
Agli albori dell'evoluzione dei mammiferi, la struttura del cervello anteriore era simile alla struttura del cervello dei moderni rappresentanti lissencefalici di roditori e lagomorfi (vedi Fig. III-18, b; III-19, a, b; III-24, a). I mammiferi arcaici cercavano cibo, partner sessuali e navigavano nello spazio usando il loro senso dell'olfatto. La maggior parte degli autori che si sono occupati dei problemi della loro evoluzione concordano con questo punto di vista (UIinski, 1986). In questa fase dell'evoluzione, i rettili antenati dei mammiferi erano privati della capacità di utilizzare gli altri sensi con la stessa efficacia dell'olfatto. Apparentemente vivevano nell'oscurità degli strati inferiori di macerie di carbonio, dove l'olfatto era il recettore distante più efficace. Oltre all'olfatto, lì potevano essere utilizzati anche l'udito e la sensibilità tattile. Il sistema visivo e la visione dei colori erano praticamente inutili e perdevano progressivamente le loro caratteristiche originarie.
I mammiferi arcaici rimasero in questo stato per molto tempo. C'era abbastanza tempo per creare connessioni integrative tra i centri corticali genitali del sistema vomeronasale e i sistemi sensomotori di altre parti del cervello. Un nuovo centro decisionale sorse sulla base del modesto rudimento corticale del sistema olfattivo sessuale rettiliano. Chiaramente originariamente comprendeva i centri vomeronasale, motorio e gustativo.
Nella prima fase dell'evoluzione dei mammiferi, il sistema uditivo fu migliorato grazie ai tubercoli posteriori del quadrigemino. Sono più sviluppati nei mammiferi che nei rettili e negli uccelli (vedi Fig. III-22, d). Pertanto, quando emerse dai labirinti del Carbonifero, il potenziale antenato dei mammiferi aveva un senso dell'olfatto sviluppato, tubercoli uditivi nella parte superiore del mesencefalo e un rudimento corticale che integrava i centri olfattivi, sessuali, motori e del gusto (Fig. .III-27, UN, B).
Sorge una domanda del tutto naturale sul destino futuro di queste creature. Di solito si presume che i piccoli antenati dei mammiferi fiutassero le loro prede nel suolo della foresta di notte e che durante il giorno si nascondessero nelle tane o tra le radici degli alberi. Questo è un presupposto del tutto corretto, sebbene spieghi solo la possibilità di sviluppare l'olfatto. Tuttavia, con questo stile di vita non si possono rilevare ulteriori stimoli per lo sviluppo della neocorteccia, e soprattutto degli emisferi cerebellari. Al contrario, gli animali scavatori crepuscolari hanno un cervelletto più che modesto. La rapida evoluzione dei centri corticali sensomotori e del cervelletto richiedeva un ambiente tridimensionale incredibilmente complesso che non era mai stato incontrato prima dai vertebrati. Si dovrebbe presumere che non sia stato il suolo, ma un altro ambiente a causare la comparsa di una sensibilità somatica sviluppata.
Nella ricerca dell'ambiente di evoluzione dei mammiferi, un aiuto significativo può essere fornito dall'analisi di un altro sistema recettoriale, che è difficile sopravvalutare: la sensibilità somatica. Il tegumento dei mammiferi ha acquisito una straordinaria serie di meccanorecettori di vario tipo. Sono specializzati nel percepire vari tipi di vibrazioni, pressione, tatto, riscaldamento e raffreddamento. Gli abitanti del suolo non hanno assolutamente bisogno di un insieme così diversificato di recettori cutanei, soprattutto perché i moderni mammiferi del suolo (ratti talpa senza pelo) hanno addirittura peli ridotti. È improbabile che un sistema somatosensoriale e peli sviluppati possano essersi formati negli animali che conducono uno stile di vita semi-sotterraneo.
Apparentemente, gli antenati rettili dei mammiferi, dopo aver lasciato le macerie del Carbonifero, si trasferirono tra le chiome degli alberi (vedi Fig. III-27, V, G). La “migrazione” verticale dai frangivento delle piante poco illuminate al mondo crepuscolare delle chiome degli alberi sembra del tutto naturale. Questa transizione non fu un cambiamento radicale nella biologia degli antenati rettili dei mammiferi. Sono stati preservati un ambiente di vita tridimensionale simile e l'importanza di un apparato vestibolare già ben sviluppato. È probabile che la transizione dai livelli più bassi di detriti forestali di carbonio alla chioma arborea sia avvenuta ripetutamente, ma con risultati diversi. Solo dopo la comparsa della specializzazione primaria del cervello del rettile secondo la tipologia olfattiva poterono emergere i presupposti necessari per la formazione del gruppo “arboreo” dei mammiferi arcaici. Nelle chiome crepuscolari degli alberi è necessario esattamente l'insieme di acquisizioni neurosensoriali, analitiche e riproduttive noto nei mammiferi moderni.
La vita dei macrosmatici nelle chiome degli alberi praticamente escludeva la riproduzione nei nidi o nelle cavità. Per i piccoli animali con un senso dell'olfatto sviluppato, la covata di uova di un altro era e rimane un alimento ideale e accessibile, quindi la viviparità, ereditata dagli antenati rettili, è stata ulteriormente sviluppata. Era necessario prolungare il più possibile lo sviluppo intrauterino dell'embrione. Ciò ha permesso di evitare la formazione di un nido e l'attaccamento a un territorio specifico. La madre si è spostata con il cucciolo in cerca di cibo, il che ha aumentato le probabilità di sopravvivenza.
Il modo più semplice per aumentare la durata dello sviluppo intrauterino è rifiutarsi di nutrire l'embrione dal tuorlo. Le riserve di tuorlo non possono essere aumentate indefinitamente nell'utero materno. È molto più efficace utilizzare un semplice scambio diffusivo di ossigeno, acqua e metaboliti tra la parete del sacco vitellino e l'utero. Apparentemente, questo metodo è stato utilizzato per risolvere il problema dello sviluppo intrauterino dei mammiferi arcaici. Gli antenati arboricoli dei mammiferi erano animali molto piccoli. Ciò ha permesso loro di far crescere gli embrioni fino a dimensioni pienamente vitali utilizzando la placenta del tuorlo. I marsupiali moderni utilizzano una strategia riproduttiva simile. Tuttavia, la placenta del tuorlo consente loro di far crescere solo un piccolo embrione, che deve essere trasferito in una sacca con le ghiandole mammarie. Poiché i mammiferi arcaici erano piccoli, probabilmente non c'era bisogno di allevare gli embrioni in sacche. Solo con l’aumento delle dimensioni degli animali potrebbero sorgere difficoltà nella crescita di embrioni di grandi dimensioni. Gli animali inferiori hanno risolto questo problema con l'aiuto di una borsa e i mammiferi superiori con l'aiuto della placenta (Jameson, 1988).
Insieme allo sviluppo di strategie riproduttive efficaci nei mammiferi arcaici, il sistema sensomotorio deve aver subito i cambiamenti più evidenti. Nelle chiome degli alberi, il carico sull'apparato vestibolare è molte volte superiore anche a quello dell'ambiente acquatico tridimensionale. Anche se il pesce fa un movimento errato mentre nuota, ciò non porta a conseguenze fatali. La dipendenza dall'acqua viene mantenuta in ogni situazione e consente di correggere gli errori del motore. Per i vertebrati protoacquatici i requisiti per il sistema sensomotorio sono molto meno critici che per gli animali che vivono sui rami degli alberi e non possono volare. Gli errori sensomotori sui rami degli alberi possono avere conseguenze fatali. La gravità del pianeta è diventata un crudele esaminatore per i rettili che si sono spostati dalle macerie di carbonio alla parte superiore della foresta. Ha inoltre imposto un limite alla dimensione corporea degli antenati dei mammiferi. Gli animali di grandi dimensioni semplicemente non potevano sopravvivere agli errori di sviluppare un apparato vestibolare e un sistema sensomotorio perfetti. La caduta di animali di grandi dimensioni da un'altezza significativa porta quasi sempre alla morte o a danni non compensati, quindi la dimensione lineare degli antenati dei mammiferi non poteva superare diverse decine di centimetri. Un animale piccolo e agile doveva acquisire rapidamente non solo un perfetto apparato vestibolare, ma sviluppare anche una sensibilità somatica. Questo complesso sensoriale è ampiamente rappresentato negli emisferi cerebellari e nella neocorteccia dei mammiferi.
Tra i recettori del tegumento esistono recettori adatti a vari tipi di vibrazione. Sono emersi sistemi speciali con tempi di adattamento diversi per percepire le fluttuazioni. Recettori di vibrazione così diversi e specializzati nella pelle sarebbero completamente inutili se gli antenati dei vertebrati cercassero le prede sul terreno e nei rifiuti di foglie cadute. Al contrario, i rami e i tronchi degli alberi trasmettono perfettamente qualsiasi vibrazione. Queste vibrazioni possono contenere informazioni sulla preda, su un animale del sesso opposto o sull'avvicinarsi di un pericoloso predatore. Tali segnali dovevano essere differenziati dalle vibrazioni sicure ma varie degli alberi stessi, in modo che lo sviluppo della sensibilità somatica nei rettili arboricoli fosse biologicamente completamente giustificato. Nella prima fase dell'evoluzione dei rettili antenati dei mammiferi, la sensibilità dei meccanocettori del tegumento potrebbe essere stata lungi dall'essere perfetta come negli animali moderni. Questa carenza potrebbe essere compensata dallo sviluppo di formazioni sensibili specializzate. Tuttavia, complessi recettori incapsulati come quelli terminali di Ruffini, Pacini, Meissner o Krause non potevano attivarsi istantaneamente per svolgere le loro funzioni specializzate.
Apparentemente, nella prima fase dello sviluppo della sensibilità somatica, venivano utilizzate terminazioni nervose libere, ben sviluppate in tutti i vertebrati. La difficoltà è che le terminazioni nervose libere hanno capacità sensoriali limitate. Un semplice aumento del loro numero nel derma non sarebbe stato in grado di risolvere i complessi problemi somatosensoriali degli antenati arboricoli dei mammiferi.
L'aumento della meccanosensibilità somatica è stato ottenuto utilizzando i capelli (Spearman e Riley, 1980). I capelli sono diventati una sorta di amplificatore di segnale meccanico. In effetti, il modo più semplice per amplificare un segnale meccanico è creare una leva di Archimede a bracci disuguali. Il braccio lungo diventerà un rilevatore meccanico e il braccio corto diventerà un recettore associato ad una terminazione nervosa libera. È chiaro che la sensibilità di un tale sistema sarà determinata dalla forma, dimensione e massa della leva, dalla sua rigidità e dalla sensibilità della terminazione nervosa. Se ci sono molti di questi recettori, sarà garantita la differenziazione delle informazioni somatiche in direzione, forza e frequenza. È possibile che lo sviluppo di un sistema di recettori somatici così specializzato abbia portato alla comparsa di peli recettoriali (Hudspeth, 1985). Successivamente venne utilizzato per conservare il calore, mascherando la sua funzione primaria. L'origine dei recettori dei capelli è indicata anche dallo sviluppo del loro sistema muscolare. La regolazione fine dello scambio di calore può essere effettuata in altri modi fisiologici, ma non esiste altro modo per modificare dinamicamente la sensibilità dei meccanorecettori che intrecciano il follicolo pilifero, quindi in caso di pericolo il pelo di molti animali si rizza di riflesso. Ciò aumenta la meccanosensibilità dell'attaccatura dei capelli come risultato della tensione della “leva” del recettore.
In un lontano passato, la tensione nei peli recettoriali degli antenati dei mammiferi aumentava la precisione delle informazioni somatosensoriali. Ciò ha permesso di scegliere una forma di comportamento adeguata in risposta alla situazione che si è venuta a creare. A giudicare dal supporto neurologico, questo meccanismo di aumento della sensibilità somatica è sorto agli albori dell'evoluzione dei mammiferi. È sopravvissuto fino ad oggi come reazione involontaria a qualsiasi eccitazione inaspettata. Di conseguenza, la sensibilità somatica primaria dei rettili antenati dei mammiferi si è sviluppata sulla base delle terminazioni nervose libere associate ai peli recettoriali. Una prova indiretta a favore di questo punto di vista è l’elevata innervazione dei fusti dei capelli e dei follicoli piliferi. In alcuni animali, attorno alla base del pelo possono essere raggruppate fino a 20 fibre nervose sensoriali. Questo sistema meccanorecettore ha la soglia di eccitazione più bassa ed è sensibile alle vibrazioni con una frequenza di circa 35 Hz.
Fornendo un aumento della sensibilità somatica nel modo più primitivo, gli antenati dei mammiferi gettarono le basi per l'evoluzione a lungo termine di recettori incapsulati perfetti. Diventeranno più efficaci delle terminazioni nervose libere e delle terminazioni associate solo dopo milioni di anni. Un sottoprodotto della formazione del sistema somatosensoriale primario erano i capelli primitivi. Il suo ulteriore sviluppo come strato di isolamento termico apparentemente è avvenuto molto più tardi rispetto alla formazione delle funzioni meccanosensoriali.
Parallelamente all'apparato sensoriale periferico si svilupparono meccanismi centrali per l'analisi dei segnali somatici e propriocettivi. Sono la sensibilità somatica e il sistema motorio ad essere rappresentati da estesi campi nella neocorteccia dei mammiferi lissencefalici (vedi Fig. III-24). Apparentemente, la necessità di sviluppare il controllo corticale su questi due sistemi è diventata una delle ragioni principali dell'evoluzione del prosencefalo. Ciò è indicato dallo sviluppo parallelo del neostriato (gangli della base) dei mammiferi. Neoplasie così estese e specializzate nella parte ventrale del prosencefalo non si sono mai verificate in altri vertebrati (Reiner, Brauth, Karten, 1984). È interessante notare che questi enormi centri nucleari elaborano informazioni sensomotorie e cinestetiche che provengono da altre parti del cervello. Alleviano la corteccia sensomotoria dal controllo dei movimenti involontari.
Va sottolineato che, parallelamente all'espansione della rappresentazione dei recettori somatici nella neocorteccia, si sono formate connessioni simili dei recettori cutanei con gli emisferi cerebellari. Gli emisferi cerebellari accoppiati si trovano solo nei mammiferi a causa dello sviluppo di una sensibilità somatica fenomenale e della coordinazione di movimenti complessi. Questo sviluppo del cervelletto non può essere associato ad alcuna condizione standard nella storia dei vertebrati prima dei mammiferi. Perfino l’ambiente acquatico tridimensionale in cui i vertebrati primordiali si sono evoluti per centinaia di milioni di anni non è riuscito a portare i loro sistemi sensomotori allo stesso livello di sviluppo di quelli dei mammiferi.
Il cervelletto “mammifero” dei triconodonti si è formato in soli 30-40 milioni di anni. Il motivo della sua comparsa va ricercato nelle chiome degli alberi ad alto fusto, dove la vita di qualsiasi animale dipende dall'efficacia dell'analisi dei segnali somatici e dalla coordinazione dei movimenti dell'intero corpo. Nei mammiferi, l'intera superficie del cervelletto è occupata da una corteccia organizzata in modo complesso, costituita da neuroni specializzati. Ciascuna superficie recettoriale del corpo è rappresentata da un'area rigorosamente definita della corteccia cerebellare. Ciò ha comportato un aumento della superficie delle strutture corticali del cervelletto dei mammiferi migliaia di volte rispetto al cervelletto dei rettili. Infatti, come risultato dell'espansione laterale, apparvero emisferi cerebellari accoppiati. Una conseguenza dello sviluppo delle connessioni cerebellari interemisferiche è stata la formazione del ponte rombencefalo dei mammiferi, che non è presente nei rettili e negli uccelli. Il motivo della formazione del ponte era la necessità di un costante confronto operativo delle informazioni somatiche provenienti dalla metà destra e sinistra del corpo e della correzione motoria della posizione del corpo. La sopravvivenza dei mammiferi arcaici nelle chiome degli alberi dipendeva direttamente dallo sviluppo dell'apparato analitico della sensibilità somatica e sensomotoria. Il cervelletto divenne una sorta di automa cinestetico che integrava segnali somatici, sensomotori e vestibolari. Svolgendo queste funzioni, permetteva agli antenati dei mammiferi di risolvere inconsciamente problemi di movimento in un ambiente tridimensionale complesso.
L'evoluzione dei mammiferi arcaici nelle chiome degli alberi aiuta a spiegare lo sviluppo specifico di altri organi di senso e la loro rappresentazione cerebrale. Il complesso ambiente tridimensionale richiedeva modi completamente nuovi di valutare lo spazio circostante partendo dalla visione semplificata degli antenati dei mammiferi. Era necessario non solo vedere l'oggetto, ma determinare con estrema precisione la distanza e valutarne le proprietà. Valutare erroneamente la distanza da un ramo nella chioma di un albero di solito costa la vita. La visione binoculare e la rappresentazione corticale di questo sistema nel prosencefalo sono del tutto giustificate.
Va sottolineato che la vista, la sensibilità somatica, la propriocezione e l'apparato vestibolare dell'orecchio interno sono i principali input sensoriali ai nuclei vestibolari del tronco encefalico. L'integrazione di questi segnali consente ai mammiferi di posizionare il proprio corpo nello spazio e di controllare la precisione dei propri movimenti. I nuclei vestibolari dei mammiferi sono una formazione unica. Sono molto più sviluppati di quelli dei rettili e degli uccelli. Apparentemente, un tale sistema multifunzionale di controllo vestibolare e cinestetico poteva svilupparsi solo nelle dure condizioni delle chiome degli alberi. In un tale ambiente c'erano tutte le condizioni per la formazione di un sistema uditivo unico nei mammiferi. L'orecchio esterno, che può essere orientato verso una sorgente sonora, potrebbe essere sorto nel complesso ambiente acustico delle chiome degli alberi. I moderni mammiferi arboricoli hanno proprio questi gusci uditivi esterni. Avendo acquisito le caratteristiche strutturali elencate del sistema nervoso nelle chiome degli alberi, i mammiferi “scesero” ripetutamente a terra. I monotremi furono i primi a ritornare all'esistenza ultraterrena (vedi Fig. 111-27, cf), poi i marsupiali e, più tardi, i mammiferi placentati (vedi Fig. III-27, e-m). Apparentemente pipistrelli e primati erano completamente formati tra le chiome degli alberi. Il passaggio dei primati all'esistenza terrestre fu il primo passo verso l'emergere dell'uomo.
L'acquisizione più importante del cervello dei mammiferi che vivevano tra le chiome degli alberi era la capacità di prevedere gli eventi. La capacità di prevedere un evento, il risultato di un movimento, le conseguenze di una caccia o di un conflitto intraspecifico contraddistingue anche i mammiferi moderni. La capacità del sistema nervoso di prevedere l'esito di un'azione non ancora compiuta era assente in altri vertebrati. I mammiferi hanno pagato a caro prezzo questa capacità con errori commessi lontano dalla terraferma. Scesi sulla terra per la seconda volta, i mammiferi possedevano non solo centri associativi di tipo rettile, ma anche una modesta capacità di valutare i risultati di azioni immediate. Questa acquisizione funzionale dei mammiferi si basa sulla sovrabbondanza di neuroni e connessioni che si sono formate nella neocorteccia. Solo l'eccesso di memoria e l'esperienza individuale hanno permesso ai mammiferi di occupare una posizione dominante nel mondo animale.
Teoria dei mezzi di transizione
L'evoluzione del sistema nervoso dei vertebrati si basa su modelli morfologici generali. Si riducono a cambiamenti quantitativi e qualitativi nel sistema nervoso centrale e periferico. Tuttavia, a differenza di altri sistemi corporei, qualsiasi cambiamento strutturale provoca una profonda ristrutturazione del comportamento. Il risultato è un cambiamento nelle forme di interazione dell'organismo con l'ambiente esterno. Le nuove proprietà morfofunzionali del sistema nervoso non sempre portano a risultati positivi. Alcune di queste proprietà sono diventate la base per la prosperità a breve termine di un gruppo o una specializzazione senza uscita, altre hanno dato ai vertebrati l'opportunità di padroneggiare risorse infinite e hanno aperto promettenti percorsi di evoluzione. Nella storia naturale del sistema nervoso ci sono state e rimangono decisioni morfologiche che condannano i loro proprietari sia all'inevitabile estinzione che alla prosperità. La maggior parte degli animali moderni sono esempi di adattamenti più o meno riusciti, ma senza uscita. La loro scomparsa era predeterminata al momento dell'inizio della specializzazione strutturale del sistema nervoso.
Il sistema nervoso ha una proprietà notevole: può modificare quasi istantaneamente il comportamento di un animale e l'attività fisiologica dei suoi organi, per poi ripristinare altrettanto rapidamente la situazione originale. La reversibilità dei cambiamenti rapidi e assolutamente necessari lo rende uno strumento inestimabile nel mondo biologico. Tuttavia, la gamma delle possibili ristrutturazioni del sistema nervoso è limitata dalla sua struttura. Il cervello può fornire solo quell'insieme di decisioni istintive o associative fornite dai sistemi sensomotori del corpo. L'orso non agiterà le zampe, anche se può effettivamente volare. Solo una persona civilizzata decide facilmente di compiere tali azioni, poiché il suo cervello ha quasi perso il contatto con il mondo reale del pianeta. In altre parole, tutti i vertebrati sono prigionieri del passato evolutivo del loro sistema nervoso. Consentendo agli animali di adattarsi rapidamente ai cambiamenti momentanei dell'ambiente, il cervello forma una sorta di struttura nascosta per i massimi cambiamenti possibili. Sono queste strutture che predeterminano i limiti dei cambiamenti adattativi reversibili nel comportamento di una particolare specie.
L'evoluzione morfologica del sistema nervoso è necessaria come strumento per espandere i confini delle capacità adattative. I cambiamenti strutturali nel cervello rimuovono le restrizioni da alcune reazioni comportamentali e ne modellano altre. Questo processo può continuare finché il cervello non diventa troppo specializzato per ulteriori cambiamenti. Tuttavia, i cambiamenti quantitativi o qualitativi nel sistema nervoso rimangono l’unico modo per superare le capacità standard. Va sottolineato che i cambiamenti quantitativi nel sistema nervoso possono avvenire molto più velocemente di quelli qualitativi. Sono la risorsa primaria per gli adattamenti strutturali del tessuto nervoso. I cambiamenti morfologici qualitativi nel cervello sono estremamente difficili e di solito richiedono condizioni speciali o molto tempo. Questa differenza tra cambiamenti quantitativi e qualitativi nelle strutture neurali è mediata dalla posizione speciale del cervello e del midollo spinale nel sistema di organi e tessuti dei vertebrati.
Il sistema nervoso centrale non è coinvolto nelle interazioni biomeccaniche del corpo con l'ambiente esterno. Ciò non significa che quando i muscoli si contraggono, le articolazioni si muovono, la pelle si deforma o il cibo attraversa l’intestino, i nervi non sono soggetti a stress meccanico. Hanno resistenza e duttilità conosciute e possono sopportare carichi piccoli e a breve termine. Tuttavia, non stiamo parlando delle proprietà meccaniche del sistema nervoso. Al contrario, ciò che è più interessante per la morfologia evolutiva è che il sistema nervoso è estremamente protetto da qualsiasi stress, ad eccezione dei meccanorecettori specializzati. L'intera evoluzione del cervello e del midollo spinale avviene all'interno del cranio e sotto la protezione degli archi neurali delle vertebre. Sono separati dagli elementi scheletrici da tre meningi e dal liquido cerebrospinale. Tuttavia, la forma del sistema nervoso centrale varia leggermente a seconda dello scheletro. Basti pensare che nel periodo embrionale dello sviluppo sono il cervello e il midollo spinale gli induttori della differenziazione scheletrica, e non viceversa. È più giusto dire che la forma del cranio e degli archi neurali delle vertebre è secondaria rispetto all'anatomia del sistema nervoso centrale. Di conseguenza, il cervello e il midollo spinale cambiano forma indipendentemente da eventuali trasformazioni biomeccaniche nel corpo dell’animale.
Questa posizione speciale del cervello contiene un enorme potenziale per qualsiasi trasformazione strutturale. La vera fonte dei cambiamenti quantitativi è la variabilità individuale stabile del sistema nervoso. Studi speciali condotti su anfibi, rettili e mammiferi hanno dimostrato che nelle larve vitali o negli animali giovani ottenuti da una coppia di genitori, una variabilità quantitativa nel cervello del 20-22% è standard. I neuroni sono stati contati sia in tutte le parti del cervello che nei principali analizzatori periferici. È stata riscontrata una certa variazione nei valori di variabilità a seconda della regione del cervello. Le strutture più antiche (romboencefalo e midollo allungato) erano caratterizzate da una variabilità del 7-13% e per quelle evolutivamente nuove - 18-25%. Tuttavia, la variabilità quantitativa copriva quasi tutte le parti del cervello. I limiti identificati di variabilità sono stati stabiliti in un gruppo geneticamente omogeneo di animali, discendenti di una sola coppia di genitori.
Se usiamo una persona per valutare la variabilità del sistema nervoso, allora una doppia differenza nella massa cerebrale non esaurirà tutte le varianti presenti e vitali (Savelyev, 1996), quindi la conclusione più obiettiva riguarderà il normale intraspecifico 15-25 % variabilità del cervello. Ciò significa che un animale è costantemente diverso dall'altro per una certa quantità di tessuto nervoso. Per l'anamnia, questa risorsa può variare da diverse migliaia a decine di milioni di neuroni, e per gli amnioti, da centinaia di migliaia a diversi miliardi di cellule. Considerando che ogni neurone ha molti contatti con altre cellule e può essere portatore di memoria, possiamo supporre una notevole differenza nel comportamento degli individui anche nella popolazione più omogenea. Le prove etologiche dell'individualizzazione del comportamento sono numerose e coprono quasi tutti i gruppi di vertebrati. Ciò significa che in ogni popolazione ci sono individui che riescono a risolvere certi problemi meglio o peggio di altri. Se la situazione biologica è stabile, nessuno trarrà mai vantaggio da questa differenza nelle potenziali capacità del cervello.
Le differenze quantitative nel cervello diventano significative quando l’ambiente è instabile, c’è un’elevata competizione sessuale o c’è una risorsa alimentare chiara ma inaccessibile. Si rivelano una riserva decisiva quando gli insiemi istintivo-associativi di forme di comportamento specie-specifiche sono completamente esauriti. Se una forma di comportamento individualizzata fornisce un notevole guadagno nell'accesso al cibo, allora è rafforzata dai successivi vantaggi riproduttivi e aumenta la probabilità di preservare le caratteristiche quantitative del cervello di un dato individuo. Apparentemente, è questo meccanismo alla base dei cambiamenti adattativi su larga scala nel cervello della maggior parte dei vertebrati protozoi. A seconda del tipo di alimentazione e dello sviluppo degli organi di senso, il loro cervello aumenta in modo differenziale di dimensioni (vedi § 27). Questo percorso nell'evoluzione del sistema nervoso è efficace per risolvere particolari problemi di adattamento nel quadro della struttura esistente del sistema nervoso. I principali eventi evolutivi che portano a un cambiamento nell'habitat e all'emergere di nuovi taxa sistematici di ordini elevati richiedono cambiamenti qualitativi nel sistema nervoso.
La comparsa di strutture qualitativamente nuove nel sistema nervoso richiede molto tempo e condizioni molto speciali. Queste condizioni devono essere diverse dall'habitat tradizionale ed esercitare un'attrazione irresistibile per i vertebrati. Le garanzie di tale attrattiva sono il cibo abbondante e la riproduzione di successo. Se un ambiente così biologicamente benefico viene mantenuto per lungo tempo, gli animali hanno la possibilità di acquisire una struttura neuromorfologica qualitativamente nuova.
Poche condizioni ecologiche di questo tipo si sono verificate nella storia dei vertebrati e tutte sono state contrassegnate dalla comparsa di animali con strutture del sistema nervoso qualitativamente nuove. Il primo evento di questo tipo fu l'emergere dei cordati. Come descritto sopra, la comparsa dei cordati fu un evento piuttosto casuale e non un modello evolutivo fatale (vedi § 26). Un gruppo di piccoli vermi piatti simili ai turbellari continuava ad abitare acque poco profonde e ricche di cibo. Essendo filtratori e conducendo uno stile di vita passivo, queste creature simili a vermi piatti hanno cercato di prendere piede nei territori alimentari più redditizi. Per fare ciò, immergevano la parte posteriore del corpo nei sedimenti del fondo. Tale ancoraggio è diffuso tra i moderni invertebrati bentonici. Le conseguenze a lungo termine di queste semplici azioni adattative degli antichi vermi furono il cordone nervoso dorsale e la notocorda muscolare, che ne impedirono la deformazione. L'essenza dei cambiamenti qualitativi nel sistema nervoso a due o quattro catene degli antenati dei cordati simili a vermi era costituita da diversi eventi successivi. Nella versione a doppio filamento, il verme ruotava di 90 gradi rispetto a una delle superfici laterali del corpo. Con una struttura a quattro catene del sistema nervoso, si nota una fusione di catene nervose dorsali e ventrali accoppiate. In entrambi i casi, una ristrutturazione qualitativa del sistema nervoso è culminata nella fusione rostrocaudale dei gangli segmentali della catena nervosa dorsale con la successiva formazione del ventricolo centrale. Parallelamente si ebbe una divisione dei nodi della catena nervosa ventrale fino al livello dei gangli somatici (vedi § 26). Sono diventati la base per l'innervazione degli organi interni. I cordati non sarebbero comparsi senza uno specifico ambiente di transizione. La profondità dell'acqua bassa, l'abbondanza di cibo e le condizioni adatte alla riproduzione garantivano la prosperità di qualsiasi filtro di fondo. Tra le tante opzioni di adattamento a un ambiente così favorevole, l'emergere del morfotipo cordato è stata solo una delle opzioni di successo. In questa situazione ha giocato un ruolo decisivo un ambiente ricco di cibo, che è diventato stimolo per cambiamenti morfologici in molte specie. L'ulteriore evoluzione dei cordati ebbe luogo in condizioni più diverse e portò all'emergere dell'intera diversità dei vertebrati protoacquatici (vedi § 29).
Il cervello ha subito il secondo cambiamento qualitativo fondamentale dopo che i vertebrati hanno raggiunto la terra. Questo evento portò a importanti cambiamenti morfologici sia nel sistema nervoso che in altri organi. Si formarono arti, respirazione polmonare, tegumenti specializzati e una serie di altre caratteristiche che consentirono ai tetrapodi arcaici di passare all'esistenza terrestre. Tali estesi riarrangiamenti morfofunzionali dell'apparato analizzatore ed effettore del sistema nervoso non potrebbero verificarsi in un breve periodo di tempo e al di fuori di uno speciale ambiente di transizione. Erano particolarmente necessari per i cambiamenti qualitativi nel sistema nervoso, poiché in termini quantitativi il cervello degli anfibi è chiaramente inferiore agli animali protoacquatici specializzati. Quando si raggiunge la terra, nel sistema nervoso degli antichi anfibi emergono il sistema olfattivo vomeronasale, il controllo della respirazione e un complesso di centri di controllo del tronco encefalico per gli arti. I sistemi visivo, uditivo e vestibolare hanno subito cambiamenti. Gli ecosistemi di transizione tra habitat acquatici e terrestri potrebbero essere peculiari labirinti del suolo o detriti di foreste di carbonio (vedi § 31). In un ambiente così transitorio, sia i movimenti natatori che il supporto delle pinne potrebbero essere utilizzati per molto tempo. Quando l'umidità dei labirinti era elevata, la respirazione cutanea, le branchie e le gemme polmonari funzionavano simultaneamente. Lo sviluppo degli organi sensoriali e dei sistemi motori acqua-aria in un ambiente di transizione era giustificato dai vantaggi biologici forniti dallo sviluppo di territori ricchi di cibo e ben protetti (vedi § 33). Apparentemente, sia i labirinti del suolo che i detriti di carbonio dei tronchi degli alberi hanno creato un ambiente di transizione unico per la graduale evoluzione del sistema nervoso degli antichi anfibi. Solo con lo sviluppo a lungo termine dei cambiamenti morfologici potrebbero comparire i centri spinali e il nucleo rosso per il controllo degli arti, l'organo vomeronasale e il bulbo olfattivo aggiuntivo, nonché i centri uditivi e vestibolari secondari.
Il terzo periodo storico nello sviluppo del sistema nervoso può essere considerato la formazione del cervello dei rettili arcaici. Il periodo rettiliano fu il più fruttuoso nella storia dei vertebrati. I rettili stabilirono i principi di base dell'evoluzione strutturale del cervello amniotico. Nei rettili si è formato per la prima volta un dipartimento associativo nel sistema nervoso. Ha avuto origine dal mesencefalo ed è stata un'acquisizione così riuscita che i rettili sono diventati il gruppo di vertebrati più dominante per milioni di anni. Il centro associativo del mesencefalo non si sarebbe mai formato senza una seria necessità biologica. È nato all'inizio dell'evoluzione dei rettili come un modo per adattarsi a un ambiente aggressivo. I rettili arcaici avevano bisogno di confrontare costantemente le informazioni provenienti da sensi diversi e prendere decisioni complesse. Le decisioni sono state guidate dal costante adattamento del comportamento a una situazione in rapido cambiamento. Il cervello dei vertebrati e degli anfibi protoacquatici non possedeva queste proprietà. Hanno scelto una delle forme di comportamento istintive secondo principi completamente diversi. La scelta degli anfibi si è basata sulla competizione tra i centri cerebrali della rappresentazione dell'analizzatore (Fig. III-28). Un semplice confronto del livello di eccitazione era una condizione sufficiente per l'attuazione di uno dei programmi istintivi. I rettili furono i primi a possedere un tipo completamente nuovo di strumento analitico (vedi Fig. III-28). Funzionava secondo il principio del confronto delle informazioni provenienti da ciascun organo di senso. Il ruolo decisivo cominciò ad essere giocato dal contenuto del segnale dell'analizzatore e non dal fatto stesso dell'eccitazione (vedi § 37). È un dato di fatto, i rettili hanno sviluppato le basi del principio associativo della ricerca di soluzioni. È chiaro che vediamo i segni più rudimentali di questa proprietà distruttiva del cervello, ma sono sorti proprio nei rettili. La storia dei rettili è stata probabilmente molto più ricca di esperimenti neurologici di quanto possiamo immaginare. Basti menzionare un'altra acquisizione storica dei rettili: le strutture corticali del prosencefalo (vedi § 39). La competizione sessuale, combinata con l'incredibile sviluppo dell'olfatto e del sistema vomeronasale dei rettili, ha fornito le basi per l'emergere delle strutture corticali. Le strutture corticali del prosencefalo si sono formate sulla base di un nuovo centro che garantisce l'integrazione dei segnali sessuali con il resto dei sensi. Questo centro di integrazione sessuale ha gareggiato per un breve periodo con il tetto associativo del mesencefalo, ma la sua attività si è manifestata solo durante la stagione riproduttiva. Apparentemente, per una riproduzione di successo, i rettili arcaici dovevano subordinare tutti i sistemi corporei a questo compito, e qualsiasi attività secondaria, inclusa la ricerca del cibo, doveva essere ignorata (Fig. III-29).
L'associazione ed i centri corticali del cervello rettiliano non avrebbero potuto formarsi senza condizioni molto particolari. Tuttavia, supponiamo che i rettili arcaici si siano semplicemente stabiliti sulla superficie della Terra. Senza una seria concorrenza da parte di anfibi, insetti e piante, diventerebbero rapidamente il gruppo dominante senza profondi cambiamenti nel sistema nervoso. In tali condizioni non è possibile presentare alcuna base reale per il suo miglioramento. Inoltre, è impossibile trovare ragioni esterne per la formazione di un senso dell'olfatto così ipertrofico da portare all'emergere di strutture corticali del prosencefalo. Di conseguenza, gli eventi reali si sono sviluppati secondo uno scenario completamente diverso e non hanno avuto nulla a che fare con l'idilliaca dispersione dei rettili sulla superficie del pianeta.
Molto probabilmente, è molto probabile un'evoluzione piuttosto lunga dei rettili arcaici in un ambiente di transizione specializzato. Questa nicchia ecologica evidentemente non era adatta alla prosperità indisturbata di un giovane gruppo di vertebrati. Molto probabilmente, tutte le acquisizioni neurologiche dei rettili sono nate come adattamenti adattivi a un habitat estremamente complesso e ad un ambiente competitivo aggressivo. Tale mezzo avrebbe potuto benissimo essere costituito da detriti di legno carbonioso provenienti da tronchi di piante (vedi § 38). Questo ambiente era in parte utilizzato anche dagli anfibi, ma evidentemente vi si recavano per procurarsi cibo abbondante e garantito. Il cibo era molto probabilmente costituito da vertebrati acquatici primordiali, che utilizzavano i detriti di carbonio come luoghi convenienti per la riproduzione. Nel corso del tempo, hanno cambiato siti di riproduzione o l'acqua si è ritirata. Quando la fonte di cibo si prosciugò per un motivo o per l'altro, gli anfibi iniziarono a essere usati come cibo per i loro simili. Ciò ha portato a una competizione senza precedenti e a una rapida selezione per le proprietà e le capacità associative del cervello.
L'ambiente di transizione per la formazione dei rettili erano i detriti vegetali di carbonio, dove l'ambiente tridimensionale imponeva maggiori esigenze al sistema vestibolare e agli analizzatori distanti. L'assenza di luce ha portato l'olfatto ad un livello di sviluppo morfofunzionale qualitativamente diverso. Era utilizzato come importante analizzatore remoto e sistema per il controllo del comportamento sessuale. Il sistema uditivo si è evoluto attivamente, il che non è meno efficace per l'orientamento al buio.

Nel corso di diverse decine di milioni di anni di feroce concorrenza nei labirinti delle piante di carbonio, si è sviluppato un cervello rettiliano unico con un insieme abbastanza perfetto di strutture neurologiche e un centro associativo efficace. Con il suo aiuto sono stati risolti i problemi relativi alla ricerca di cibo, alla competizione, alla prevenzione del pericolo, ecc .. Quando è iniziata la stagione riproduttiva, l'intero cervello ha obbedito a una nuova struttura corticale nella parete del prosencefalo. Divenne un centro di controllo specializzato per il comportamento sessuale, a differenza di qualsiasi altro vertebrato prima dei rettili. Pertanto, il cervello dei rettili arcaici divenne il sistema più perfetto per risolvere i compiti biologici più importanti di qualsiasi specie: sopravvivenza e riproduzione. Per ogni compito è apparso il proprio sistema integrativo, in grado di reindirizzare l'intero organismo rettile per risolverlo. Con una tale risorsa comportamentale, i rettili emersero dalla loro culla aggressiva e divennero molto rapidamente il gruppo dominante del pianeta.
L'emergere del cervello degli uccelli non può essere considerato un evento evolutivo fondamentale associato ad una ristrutturazione qualitativa del cervello. Probabilmente gli uccelli sarebbero dovuti scomparire subito dopo la loro comparsa. Era una specializzazione adattiva senza uscita, salvata dalla perdita dell'olfatto. Un enorme substrato neurologico del sistema olfattivo è stato ereditato dagli uccelli arcaici a causa del cambiamento delle preferenze alimentari. Passando all'alimentazione in pozze poco profonde o tramite pinne, hanno smesso di usare l'olfatto come principale sistema di afferentazione. La vista divenne il principale sistema di analisi e l'udito divenne uno aggiuntivo (vedi § 43). Quando si procuravano il cibo nell'acqua, gli uccelli arcaici si muovevano sugli arti posteriori, il che portò gradualmente ad una significativa riduzione del carico sugli arti anteriori e alla parziale rudimentazione della mano. Il ruolo dell'ambiente di transizione in questo caso è stato svolto dalle acque costiere poco profonde, ricche di cibo, che fino ad oggi sono rimaste attraenti per gli uccelli.
Anche se la ristretta specializzazione degli uccelli ne garantì la rapida estinzione, il passaggio al nuoto e alle immersioni per procurarsi il cibo portò allo sviluppo di arti anteriori simili ad ali. Apparentemente questo stadio dell'evoluzione degli uccelli ha prodotto pinguini che non hanno mai volato. Le immersioni e il nuoto con gli arti anteriori hanno creato le condizioni fisiche per lo sviluppo delle ossa cave, dei potenti muscoli pettorali, del sistema di sacche d'aria dei polmoni e del rivestimento delle piume. Apparentemente, procurarsi il cibo in acque fredde è diventato uno dei principali incentivi per acquisire il sangue caldo. Gli arti natatori simili ad ali venivano utilizzati non solo per nuotare. Gli uccelli antichi utilizzavano i movimenti di battito degli arti anteriori per una sorta di "corsa sull'acqua", che divenne una fase transitoria al volo attivo (vedi § 44).

Le ali e la copertura di piume furono formate per la caccia nell'ambiente acquatico, ma furono adattate e utilizzate per il volo. In questa situazione, l’acqua è diventata il mezzo di transizione. Ha creato tutte le condizioni necessarie per il graduale accumulo di cambiamenti nel sistema nervoso degli uccelli, quindi la comparsa delle ali e il passaggio al volo non hanno causato cambiamenti radicali nel sistema nervoso centrale (vedi § 43). In relazione alla riduzione dell'olfatto negli uccelli, si sono formati centri associativi sulla base delle strutture basali del prosencefalo. Questi centri sono rappresentati dal neo e dall'iperstriato, che sono diventati la base per la formazione del comportamento complesso degli uccelli, della memoria e dell'individualizzazione del comportamento.
I mammiferi sono un gruppo neurologicamente strano. I vantaggi del loro cervello sono sorti sulla base dello sviluppo delle funzioni integrative del sistema riproduttivo. Come accennato in precedenza, la ragione principale della comparsa delle strutture corticali nel cervello dei rettili è stato lo sviluppo dell'organo vomeronasale (di Jacobson). La sua rappresentazione centrale si formava all'esterno degli antichi nuclei olfattivi del prosencefalo. I principali centri secondari dell'olfatto vomeronasale divennero le modeste strutture corticali dei rettili (vedi § 39). Su questo substrato morfologico sorsero
integrazione del comportamento sessuale dell'intero organismo rettile. Tale controllo centralizzato ha permesso di subordinare l'intero organismo a un compito e di ottenere più efficacemente il successo nella riproduzione.
I mammiferi sono andati ben oltre i rettili. Su questa struttura morfologica riproduttiva-integrativa del prosencefalo si è formato un centro associativo di tipo completamente nuovo. Ha iniziato a svolgere le funzioni di monitoraggio del funzionamento dei sistemi sensoriali già stabiliti. I meccanismi autonomi del cervello sono rimasti al livello dei centri antichi e tutte le funzioni acquisite complesse si sono sviluppate a livello della corteccia del prosencefalo. Oltre al senso dell'olfatto e ai centri integrativi sessuali, il cervello dei mammiferi è caratterizzato dallo sviluppo del sistema sensomotorio e dai meccanismi di controllo cinestetico. Solo nei mammiferi il cervelletto formava emisferi accoppiati. Ha raggiunto proporzioni così gigantesche che la sua superficie spesso supera le dimensioni della neocorteccia. Inoltre, una parte significativa, e talvolta ampia, della stessa neocorteccia fornisce funzioni somatiche, sensomotorie e motorie.
Perché appaia una specializzazione così strana è necessario un ambiente molto originale. Gli stessi detriti vegetali carboniferi costituivano un complesso ambiente tridimensionale per i rettili, ma il loro cervelletto non raggiungeva nemmeno lo sviluppo del cervelletto degli uccelli. L'ambiente transitorio della comparsa dei mammiferi deve aver posto esigenze insolitamente elevate sull'analisi della posizione del corpo e sulla coordinazione dei movimenti. Sulla superficie della terra, solo nei rami degli alberi possono esserci requisiti così severi per il controllo cinestetico. A quanto pare, tutti i vantaggi sensomotori, olfattivi e uditivi fondamentali dei mammiferi si formavano nella chioma degli alberi. Questo ambiente transitorio può spiegare sia la comparsa della neocorteccia sia lo sviluppo della sensibilità somatica, che divenne uno dei principali organi di senso (vedi § 48).
Lo sviluppo della sensibilità somatica ha portato alla formazione di recettori nel derma-pelo. I capelli, innervati da terminazioni nervose libere, aumentavano effettivamente la sensibilità somatica e quindi provocavano la comparsa dei capelli. L'uso continuato dei capelli per la termoregolazione ne mascherava lo scopo primario. Nelle cime degli alberi è emersa per la prima volta un'esigenza del tutto nuova per il sistema nervoso (vedi § 49). Per i mammiferi arboricoli arcaici l'analisi comparativa delle informazioni provenienti dai vari sensi non era sufficiente. Questo modo di funzionare dei sistemi associativi non consentiva di prevedere gli eventi. Sulle cime degli alberi prevedere lo sviluppo degli eventi diventava una condizione decisiva sia per procurarsi il cibo che per la fondamentale conservazione della vita. Solo il volo potrebbe salvare i mammiferi da questi problemi. Tuttavia, solo i pipistrelli vi hanno fatto ricorso dopo la formazione dei principi di base della struttura del cervello dei mammiferi. Le principali conseguenze strutturali della vita sotto le chiome degli alberi includono una neocorteccia, un cervelletto biemisferico e una scarsa capacità predittiva. Questa caratteristica dei mammiferi, dopo la loro migrazione nel suolo e negli ambienti acquatici, ha creato per loro notevoli vantaggi comportamentali. La capacità di valutare i possibili eventi è diventata uno strumento a disposizione dei mammiferi per dominare il pianeta.
Tutti i profondi cambiamenti elencati nella struttura del sistema nervoso dei vertebrati sono causati dall'adattamento del cervello alla vita degli animali in un ambiente specifico. Senza un ambiente di transizione a lungo termine, non ci sarà tempo sufficiente per i cambiamenti nell’organizzazione strutturale del sistema nervoso. È troppo qualitativamente conservatore e quantitativamente plastico per trasformazioni morfologiche rapide e radicali. L'ipotesi dell'esistenza di ambienti di transizione può spiegare l'origine del cervello dei vertebrati moderni.
Dal libro Formica, Famiglia, Colonia autore Zacharov Anatolij AleksandrovichLA COMPARSIONE DI UNA COLONIA La formazione della stratificazione avviene solitamente in un determinato momento per ciascuna specie. È così che nel 1967 in una foresta di abeti rossi vicino a Mosca (impianto di legname di Solnechnogorsk) ebbe luogo la fase iniziale dell'identificazione di un nuovo formicaio nella formica pelosa della foresta. Durante
Dal libro Dieci grandi idee della scienza. Come funziona il nostro mondo. autore Atkins PeterPrologo L'emergere della comprensione Galileo indicò un punto di svolta in cui gli sforzi scientifici presero una nuova direzione, in cui gli scienziati - un termine anacronistico, ovviamente, per quel tempo - si alzarono dalle loro sedie, mettendo in dubbio la validità dei tentativi passati
Dal libro L'anello mancante di Edie MaitlandMaitland Eady L'anello mancante (L'emergere dell'uomo - 2) Il secondo libro della serie L'emergere dell'uomo parla della ricerca di fatti relativi all'anello mancante nella successiva evoluzione dell'uomo. È dedicato all'Australopithecus, secondo la credenza prevalente
Dal libro Embrioni, geni ed evoluzione di Raff Rudolf AEmersione dei segmenti La segmentazione dell'embrione di Drosophila si presenta sostanzialmente come una serie di invaginazioni laterali della banda germinale, formatesi quasi contemporaneamente allo stadio di gastrulazione. Nonostante l’apparente natura a mosaico di questo processo, è possibile dimostrarlo
Dal libro Teoria della nutrizione adeguata e trofologia [tabelle nel testo] autore Dal libro Teoria della nutrizione adeguata e trofologia [tabelle con immagini] autore Ugolev Alexander Mikhailovich9.3. L'emergere delle cellule Si presume che lo stadio dell'emergere della vita sulla Terra debba essere considerato il periodo in cui si formarono i sistemi cellulari più semplici, che divennero la cellula elementare della vita. Le informazioni relative a questo problema sono trattate nelle recensioni dedicate a
Dal libro Come è nata e si è sviluppata la vita sulla Terra autore Gremyatsky Mikhail AntonovichVI. La comparsa della vita sulla Terra Dagli esperimenti di Spallanzani e Pasteur sappiamo già che alle alte temperature la vita cessa. La maggior parte degli organismi muore già a 70-80 gradi Celsius. Ciò significa che la loro vita richiede determinate condizioni di temperatura. Richiesto per
Dal libro Cervello, mente e comportamento di Bloom Floyd EPacemaker del cervello dei mammiferi: nuclei soprachiasmatici Alla fine degli anni '60, il fisiologo Kurt Richter condusse una serie di esperimenti sui ratti, cercando di trovare le aree del cervello responsabili del ritmo. Ha distrutto aree specifiche del cervello, in più di 200 posti diversi
Dal libro La vita nelle profondità dei secoli autore Trofimov Boris AleksandrovicL'ORIGINE DELLA VITA L'origine della vita, la sua essenza, è uno dei misteri più difficili della scienza, poiché la vita è il fenomeno naturale più complesso a noi noto. Nessuno ha visto o osservato il suo verificarsi; Inoltre, né diretto né indiretto
Dal libro L'origine del cervello autore Saveliev Sergey VyacheslavovichCapitolo II. L'emergere delle cellule nervose e del cervello La ragione dell'emergere del sistema nervoso è stata la bassa velocità con cui si ottengono informazioni sul mondo esterno ed interno di un organismo con un'organizzazione non nervosa. I suoi tessuti erano costituiti da cellule con caratteristiche chimiche, elettromagnetiche e simili
Dal libro La nascita della complessità [Biologia evolutiva oggi: scoperte inaspettate e nuove domande] autore Markov Aleksandr Vladimirovich§ 28. L'emergere di parti del cervello Il primo periodo della storia dell'origine degli antenati dei vertebrati, prima della formazione di uno scheletro ben strutturato, è piuttosto vago. Se assumiamo che le forme ancestrali dei cordati fossero creature dal corpo molle che misuravano circa 10-15 cm,
Dal libro Stato attuale della biosfera e politica ambientale autore Kolesnik Yu.A.L'emergenza e l'ereditarietà delle modifiche usando l'esempio di Metazoon Un'altra importante questione teorica che può essere considerata usando il nostro esempio di Metazoon è la questione dell'emergenza di modifiche adattative. Questo è ciò che viene chiamata più o meno significativamente la capacità del corpo (cioè,
Dal libro Antropologia e concetti di biologia autore Kurchanov Nikolaj Anatolievich4.2. L'emergere dell'organizzazione cellulare Gli scienziati hanno dimostrato che dopo l'evaporazione dell'acqua dal volume di reazione, si formano aggregati cristallini liquidi in molecole anfifiliche simili ai lipidi e lipidiche, in cui le molecole sono disposte in strati periodici, come in
Dal libro dell'autoreL'emergere del genere Homo Poiché l'evoluzione degli ominidi è avvenuta in modo non uniforme e ha avuto un carattere "a mosaico", il confine tra gli antichi rappresentanti del genere Homo e gli australopitechi è molto labile. I criteri condizionali per l'assegnazione al genere Homo sono il volume del cervello (almeno
Dal libro dell'autoreL'emergere e l'evoluzione dell'uomo moderno L'origine dell'uomo moderno è il mistero più intrigante dell'antropogenesi. In antropologia, tutte le forme di esseri umani moderni hanno ricevuto il nome operativo “sapiens”. La maggior parte dei loro rappresentanti lo erano
Dal libro dell'autoreCaratteristiche dell'evoluzione del cervello dei mammiferi e degli esseri umani I tassi eccezionalmente elevati di evoluzione del cervello dei mammiferi, e in particolare degli umani, non hanno ancora un'unica spiegazione. Inoltre, i portatori di cervelli grandi sono solitamente caratterizzati da dimensioni corporee significative, grandi
Evoluzione del cervello nei mammiferi
L'evoluzione del cervello è caratterizzata da un progressivo aumento della sua massa, volume e complessità dell'organizzazione della neocorteccia. Durante l'evoluzione, l'archi- e la paleocorteccia vengono spinte verso il setto cerebrale.
Negli animali di diversi ordini si può osservare la graduale formazione di pieghe.
I mammiferi sono caratterizzati da diversi tipi di organizzazione cerebrale.
Tipo lissencefalico L'organizzazione del cervello è caratteristica dei monotremi (ornitorinco). Con questo tipo di organizzazione si esprime costantemente il solco centrale, che divide le strutture corticali nelle parti frontale e parietale. La paleocorteccia domina quantitativamente, mentre la neocorteccia occupa un'area relativamente più piccola delle strutture corticali. Una caratteristica distintiva di questo tipo di organizzazione è l'assenza del corpo calloso. Nel cervello lissencefalico esiste una sovrapposizione tra i campi corticali delle proiezioni di diversi tipi di sentimenti. I neuroni si sovrappongono e ricevono informazioni da canali diversi. La corteccia si sovrappone alle aree motorie e sensoriali. Nel diencefalo si approfondisce la differenziazione dei nuclei talamici: si osserva un gruppo che negli insettivori forma un gruppo associativo di nuclei. Nella corteccia c'è una regione di α-motoneuroni che regolano gli α-motoneuroni del midollo spinale, questo porta alla formazione del neocervelletto nel cervelletto, che è coinvolto nella regolazione dell'attività motoria.
I marsupiali mantengono la stessa organizzazione cerebrale.
Tipo girencefalico L'organizzazione del cervello è caratteristica degli insettivori e dei roditori. Con questo tipo di organizzazione appare il corpo calloso, aumenta l'area della nuova corteccia e compaiono solchi e convoluzioni.
Negli insettivori si formano due zone polisensoriali, analoghi delle aree associative.
Nei roditori le regioni frontale e parietale sono morfologicamente distinte, ma non fungono ancora da zone associative. Questa funzione è eseguita da diversi campi della neocorteccia di diversa localizzazione. Anche in questo gruppo di animali esiste una chiara specializzazione delle zone sensoriali per certi tipi di sentimenti, ad es. non si sovrappongono più. Nei roditori appare un'altra variante della proiezione di informazioni visive (inizialmente, l'interruttore principale era retino-tettale (monotremi, marsupiali, insettivori)) - retino-talamico. Differisce dal fatto che la selezione e l'elaborazione principale delle informazioni vengono effettuate dai corpi genicolati del talamo. Nei roditori si osservano connessioni talamo-corticali già formate, tuttavia la loro differenziazione funzionale è relativamente debole.
Tipo deuteroirencefalico L'organizzazione del cervello è caratteristica dei mammiferi carnivori. Nei predatori, oltre ai solchi e alle convoluzioni costanti, si osserva uno sviluppo significativo del ripiegamento della neocorteccia. Le zone differenziate sono chiaramente distinte: zone di proiezione speciali (informazioni di processo dal sistema sensoriale), zone motorie (motrici), zone associative ben formate, che sono rappresentate nel lobo parietale e nel lobo frontale. Le zone associative corticali fanno parte di due sistemi associativi talamo-corticali:
- Il sistema talamo-parietale è responsabile dei componenti del sistema nervoso superiore: partecipa alla formazione dei riflessi di orientamento, forma l'idea della disposizione spaziale del corpo e partecipa ai meccanismi della memoria a breve termine.
- Il sistema talamofrontale è collegato alle strutture del sistema limbico. Forma un accettore di azioni (confronta a livello delle forme corticali in che misura il risultato corrisponde all'adattamento).
Tipo ominide L'organizzazione del cervello è caratterizzata nei primati superiori da una neocorteccia a sei strati. Tutte le zone funzionali della corteccia non si sovrappongono tra loro; formano tre sistemi integrativi:
- Talamo-parietale, comprende i campi 5 e 7 della corteccia associativa e svolge le seguenti funzioni: innescare una reazione di orientamento, formare la memoria a breve termine e percepire la disposizione spaziale del corpo.
- Il sistema talamo-frontale fornisce una valutazione della situazione: un accettore del risultato di un'azione, fornisce un meccanismo per la memoria a lungo termine, la formazione di sottili connessioni temporanee come risultato di azioni comportamentali (forme più elevate di inibizione).
- Il sistema integrale talamo-temporale fornisce una maggiore regolazione delle funzioni vegetative dalla corteccia attraverso i centri integrativi del sistema nervoso centrale, principalmente l’ipotalamo, il complesso dell’amigdala e i gangli della base.
Bibliografia:
1. Nozdrachev d.C. Gli inizi della fisiologia / A.D. Nozdrachev, Yu.I. Bazhenov, I.A. Barannikov e altri - San Pietroburgo: Casa editrice Lan, 2002, -1088 p.
2. Schmidt-Nielson K. Fisiologia degli animali: adattamento e ambiente. Libro 1.- M: Mir, 1982, -416 p.
3.Fisiologia evolutiva. Parte 1 / Ed. acad. E.M.Krebs. Nella serie: "Guida alla fisiologia" - L: "Scienza", 1979, -603p.
Il cervello dei mammiferi di diversi ordini presenta diversi stadi di differenziazione della struttura interna, che riflettono gli stadi dell'evoluzione dell'attività nervosa superiore che i mammiferi hanno attraversato nel loro sviluppo storico.
Nelle cloacali, il cervello proviene da
È caratterizzato solo dallo stadio iniziale di sviluppo della volta midollare secondaria: il neopallio occupa solo una piccola parte del tetto del prosencefalo. Nei marsupiali, la sezione olfattiva del prosencefalo costituisce anche le sezioni inferiore e laterale degli emisferi, e solo il tetto è rappresentato da una nuova volta: il neopallio. Lo stadio successivo è il cervello degli insettivori e dei pipistrelli, in cui la corteccia grigia degli emisferi forma già una volta cerebrale completa e la volta primaria (archipallium) viene spostata sulla superficie mediale degli emisferi sotto forma di ippocampo. Le commissure del neopallio hanno la forma rudimentale di un anello di percorsi: il corpo calloso (corpo calloso) e il fornice (fornice). Nella maggior parte dei mammiferi placentari, i recettori olfattivi, come accennato in precedenza, svolgono un ruolo di primo piano nella loro biologia, a differenza degli uccelli, in cui la visione gioca un ruolo di primo piano. Pertanto, i centri olfattivi nel cervello, concentrati nei lobi olfattivi e nella parte inferiore del prosencefalo (sistema striato), sono molto sviluppati.Anche le percezioni visive e uditive raggiungono un alto grado di sviluppo, i cui centri sono concentrati nel mesencefalo nella regione quadrigeminale. Ma oltre a ciò, nella corteccia grigia degli emisferi si sviluppano centri cerebrali associativi superiori secondari della vista e dell'udito (lobi occipitali e temporali degli emisferi). La funzione motoria degli arti raggiunge un alto grado di sviluppo nei mammiferi, così come negli uccelli, quindi nella corteccia cerebrale sono isolati speciali centri motori secondari.
Questi centri associativi secondari sono associati al potente sviluppo nel cervello dei mammiferi di nuove sezioni del cervelletto: gli emisferi cerebellari, collegati dalla commissura trasversa del ponte.
Il cervello si è sviluppato in modo indipendente in numerosi mammiferi: dagli insetti ai primati e agli esseri umani. In questa serie, dai mezzi lemuri ai lemuri e ai primati, si osserva un indebolimento dei centri olfattivi, poiché in tutte le forme arboree la visione e l'udito stereoscopici acquisiscono un'importanza di primo piano in biologia. Un'altra caratteristica importante della trasformazione del cervello dei mammiferi è l'acquisizione del ripiegamento della corteccia cerebrale sotto forma di solchi e lobuli della corteccia grigia. I mammiferi inferiori - insettivori, roditori, lagomorfi, pipistrelli - hanno un cervello liscio. Negli animali superiori, un sistema di solchi cerebrali appare sugli emisferi del prosencefalo e sul cervelletto. La piegatura (lobi e solchi) raggiunge il suo massimo sviluppo nel cervello umano, dove nella corteccia sono concentrati tutti i centri associativi secondari di attività nervosa superiore. Il ripiegamento corticale si sviluppa rapidamente anche nei mammiferi acquatici (pinnipedi e cetacei) in connessione con nuove funzioni dell'attività cerebrale nell'acqua.
Tutti hanno cranico Negli animali, l'estremità anteriore del tubo cerebrale viene trasformata nel cervello. Nelle prime fasi dello sviluppo embrionale, il cervello lampreda rappresentato da tre ispessimenti delle pareti dell'estremità anteriore del tubo cerebrale, da cui successivamente si sviluppano tre parti del cervello: anteriore, media e posteriore, o romboidale. Il proencefalo - prosencefalo - è in connessione con l'organo olfattivo ed è, quindi, il cervello olfattivo; il mesencefalo - mesencefalo - è correlato all'organo della vista e, quindi, è funzionalmente il cervello visivo e, infine, il rombencefalo, o cervello romboidale - rombencefalo - è associato allo sviluppo degli organi VlIl e X paia di nervi cranici, cioè con gli organi dell'equilibrio e l'organo della linea laterale, nonché con gli organi interni, per cui è il centro motorio più alto, che coordina l'intero apparato motorio attraverso il midollo spinale e controlla il lavoro degli organi interni. In tutti gli altri animali cranici superiori, il cervello embrionale è rappresentato da un'estensione a forma di vescica del tubo neurale situata davanti alla notocorda. Questa vescicola precordale è chiamata vescicola cerebrale primaria - archencefalo (Fig. 156-7). La piega ventrale - plica ventralis (13) - si separa presto dalla vescicola cerebrale secondaria - deaterencefalo (12), situata epicordalmente (sopra la notocorda).

Contemporaneamente alla comparsa della piega ventrale, a causa della flessione ventrale del mesencefalo precordale, il terzo mesencefalo, o mesencefalo, si separa dal mesencefalo secondario (Fig. 157, gps); La vescicola precordale è chiamata proencefalo - prosencefalo (pc), e la vescicola epicordale - rombencefalo - rombencefalo (rh). Dalla superficie dorsale, tutte e tre le parti del cervello sono delimitate abbastanza nettamente l'una dall'altra da commissure trasversali o commissure di fibre nervose, sia davanti che dietro il mesencefalo.
Successivamente, dal proencefalo su entrambi i lati della placca terminale (al posto del neuroporo) nasce sotto forma di una vescicola cerebrale accoppiata: il telencefalo (B-9), e la restante parte del prosencefalo forma il diencefalo (10 ). Quasi contemporaneamente, il rombencefalo (metencefalo) viene rilasciato dal rombencefalo, da cui successivamente si sviluppa il cervelletto, e nei mammiferi, inoltre, il ponte e la massa principale del rombencefalo forma il midollo allungato (mielencefalo), o midollo allungato.
Le cavità delle tre vescicole cerebrali primarie sono chiamate ventricoli cerebrali. I ventricoli cerebrali laterali accoppiati sono separati dalla cavità del telencefalo (Fig. 158-I e II); la cavità del diencefalo è chiamata terzo ventricolo cerebrale (III). Comunica con quelli laterali attraverso i forami interventricolari, o di Monroe (8). In seguito alla crescita delle pareti della vescica cerebrale, il ventricolo del mesencefalo si trasforma in un acquedotto canale-encefalico (7); collega il terzo ventricolo cerebrale con il quarto ventricolo cerebrale (IV), quest'ultimo è situato nel cervello romboidale, tra il midollo allungato e il cervelletto, e comunica caudalmente con il canale centrale del midollo spinale.
Il cervello di un animale adulto è formato dalle vescicole cerebrali descritte; allo stesso tempo, le singole sezioni di essi crescono in modo molto significativo o, al contrario, mantengono il loro stato embrionale per tutta la vita. Il fattore immediato che determina lo sviluppo del cervello è la differenziazione degli organi di senso, soprattutto degli organi dell'olfatto, della vista e dell'udito (nei terrestri), e degli organi del movimento volontario, che è accompagnata dalla comparsa di vari centri di coordinamento con i loro percorsi di conduzione. Così, nei selaci, che guidano i loro movimenti principalmente con l'aiuto dell'olfatto, i lobi olfattivi del telencefalo raggiungono il massimo sviluppo nel cervello. Al contrario, nei pesci ossei gli organi visivi hanno un'importanza predominante e, di conseguenza, i centri visivi nel mesencefalo si sviluppano più fortemente, mentre il cervello olfattivo diventa meno complesso. Il rombencefalo (cervelletto) è più completo negli animali che si muovono velocemente, ma negli animali che si muovono lentamente è abbastanza elementare o addirittura assente.
Negli animali terrestri, il cervello diventa più complesso nella sua struttura a causa dell'emergere di nuovi centri (superiori) nel telencefalo e delle loro connessioni sia con varie parti del cervello e del midollo spinale, sia con tutti gli organi di senso (esterno- e interorettori). . Di conseguenza, nel telencefalo si sviluppa una sezione chiamata corteccia cerebrale, che è il centro più alto che coordina e controlla tutte le attività dell'organismo animale nell'ambiente esterno e tutti i processi che si verificano al suo interno, poiché tutti i riflessi che sorgono sono chiusi nel corteccia cerebrale.
Cervello di diamante
Il rombencefalo è costituito dal midollo allungato e dal romboencefalo, rappresentato dal cervelletto e dal ponte.
Midollo
Midollo allungato s. mielencefalo (Fig. 159 e 160-5) - è costruito in modo più uniforme negli animali, poiché in tutti essi partono i nervi cranici, che sono direttamente collegati all'apparato branchiale con i suoi derivati, all'organo dell'equilibrio e dell'udito, ai visceri e cuore. È una continuazione diretta verso la testa del midollo spinale, ma differisce nettamente da quest'ultimo nella sua struttura. Il midollo allungato si è formato a seguito della crescita delle pareti laterali del tubo cerebrale embrionale. Poiché le placche laterali si allontanavano l'una dall'altra e si trovavano lateralmente rispetto alle placche principali, la cavità del cervello romboidale si espanse in modo significativo e si trasformò nel quarto ventricolo cerebrale. Il fondo del ventricolo negli animali superiori assume la forma di una fossa a forma di diamante, sulla cui superficie, anche nei mammiferi adulti, il confine tra la placca laterale e quella principale è chiaramente visibile sotto forma di un solco di confine - sulcus limitans . Le placche laterali sono particolarmente significative nei pesci; formano i lobi uditivi laterali - lobus acusticolateralis - che si separano cranialmente anche nelle orecchie del cervello romboidale - auricole. Contengono i nuclei dell'VIII paio di nervi, che sono in connessione con l'organo dell'equilibrio, con tutta la muscolatura del corpo (attraverso il midollo spinale) e con il cervelletto. Di questi, il nucleo Deiters merita un'attenzione particolare poiché è il più costante negli animali (Fig. 163-1).
A causa della divergenza dei bordi dorsali delle placche laterali, la placca tegmentale è notevolmente allungata, ma anche nei vertebrati superiori adulti rimane allo stato embrionale sotto forma di placca epiteliale - lamina epithelialis - costituita da un unico strato di ependima cellule. Si fonde con i vasi della pia madre e negli animali adulti è chiamato velo midollare caudale, o plesso corioideo del quarto ventricolo cerebrale - velo midollare caudale, resp. plesso chorioideus ventriculi quarti (Fig. 157-B, 4). Contiene aperture attraverso le quali la cavità ventricolare comunica con lo spazio intratecale.
L'allontanamento delle placche laterali spiega anche la peculiare uscita dei nervi dal midollo allungato: i nervi motori si trovano più medialmente, mentre i nervi sensoriali si trovano più lateralmente. Poiché il midollo allungato si trova tra il midollo spinale e le parti anteriori del cervello, che costituiscono l'encefalo, da cui è separato da un istmo restringente (2), le vie centrali che vanno in una direzione o nell'altra dividono il suo midollo grigio in un certo numero di singoli gruppi di nuclei di cellule nervose. Alcuni nuclei sono i centri cellulari dei nervi cranici dal V all'XI, che si estendono dal midollo allungato. Altri, al contrario, fungono da centri intermedi per i percorsi che dal midollo spinale vanno al cervello o viceversa. Questi includono i nuclei dei funicoli dorsali e il centro motorio più antico, il nucleo motore del cappello, già presente nei pesci. Le fibre dei nuclei del funicolo dorsale seguono negli animali acquatici il cervelletto e il mesencefalo, e negli animali terrestri, inoltre, il diencefalo.
Per effetto delle cellule del nucleo motore del cappello, negli animali terrestri le cosiddette olive si separano: nasali (superiori) e caudali (inferiori), divenendo centri di associazione.
Le olive nasali compaiono per la prima volta negli anfibi in relazione alla differenziazione della coclea (l'organo dell'udito). Si inseriscono nel lemnisco laterale, cioè in un anello di vie intersecanti che collegano i nuclei del nervo cocleare (VIII paio) con il talamo ottico. Un fascio di queste fibre si trova superficialmente all'estremità anteriore del midollo allungato; è più sviluppato nei mammiferi ed è noto come corpo trapezoidale - corpus trapezoideum (Fig. 175-14).
Le olive caudali sono ben definite negli uccelli e soprattutto nei mammiferi; hanno una peculiare forma a ferro di cavallo ripiegato e ricevono impulsi dai nuclei del midollo dorsale, dal midollo spinale, dal talamo visivo e rimandano gli impulsi al midollo spinale e al cervelletto (Fig. 163).
Le vie motorie che originano dalla corteccia cerebrale formano fasci nei mammiferi sul midollo allungato ventrale, noti come piramidi. Raggiungono la loro dimensione massima nell'uomo.
rombencefalo
Negli animali inferiori, il romboencefalo è rappresentato solo dal cervelletto (cervelletto) (Fig. 159 e 160-4). La funzione del cervelletto è quella di mantenere l'equilibrio e il tono muscolare, inoltre, di coordinare il lavoro dei muscoli durante il movimento, quindi è del tutto naturale che raggiunga il suo massimo sviluppo negli animali che si muovono rapidamente (nuoto veloce, volo o corsa) o muoversi su due arti. Ciò determina anche i suoi cambiamenti legati all'età. Ad esempio, nei primati superiori il cervelletto è piccolo alla nascita e costituisce solo 1/16-1/18 dell'intero cervello, mentre negli adulti raggiunge 1/8-1/9 del peso del cervello. Raggiunge una maggiore complessità in termini di dettagli strutturali negli animali capaci di movimenti più differenziati, cioè nei mammiferi.
Il cervelletto nella forma primitiva è un corpo spaiato sotto forma di un piatto corto ma largo, già costruito nelle lamprede dalla materia cerebrale grigia e bianca. Poiché in molti animali la funzione del cervelletto diventa più complessa, la placca neurale cresce dalla parte anteriore a quella posteriore e allo stesso tempo forma pieghe trasversali o in entrambe le direzioni, cioè in avanti e all'indietro, come negli squali, o solo all'indietro. , come nei pesci ossei. In questo caso, la parte centrale - il corpo del cervelletto - corpus cerebelli - e i lobi laterali delle sue orecchie - auricole - associati all'organo dell'equilibrio sono separati. Le sezioni laterali del cervelletto sono conservate anche negli animali terrestri sotto il nome di flocculus, che però sono molto poco sviluppate a causa della riduzione degli organi della linea laterale degli animali acquatici. Negli animali terrestri, anche il cervelletto nella sua forma più primitiva rappresenta un piatto, ma più lungo, disteso orizzontalmente (nei serpenti) o verticalmente (nelle lucertole). E in essi, quando la funzione del cervelletto aumenta, la placca forma una piega trasversale, piegandosi in modo arcuato dorsale (in alcuni rettili); nei coccodrilli è diviso da solchi trasversali primari e secondari - sulcus anterior et posterior - in lobi anteriori, medi e posteriori. A quest'ultimo sono collegati i flocculi sopra menzionati (Fig. 161-4).
Negli uccelli e soprattutto nei mammiferi, oltre a quelli indicati, compaiono nuovi solchi trasversali pur mantenendo il tipo principale di struttura del corpo cerebellare sotto forma di placca arcuata in direzione dorsale. Grazie ai solchi, la superficie del corpo del cervelletto è punteggiata da giri trasversali, che hanno nomi speciali nei mammiferi. Una sezione sagittale del cervelletto mostra il midollo bianco, che penetra sotto forma di rami d'albero in ciascun giro; lo schema generale della sostanza bianca è chiamato albero della vita - arbor vitae (Fig. 183-5).
Nei mammiferi, nel lobo medio, oltre ai solchi trasversali, compaiono altri due solchi longitudinali, che separano la parte media spaiata - il verme - e i lobi laterali, o emisferi cerebellari - bemisphaerae cerebelli (Fig. 181-7, 8; 161-a, 5). Gli emisferi cerebellari nei monotremi sono ancora leggermente pronunciati; Raggiungono il loro massimo sviluppo nei mammiferi superiori grazie all'emergere della capacità di movimenti isolati degli arti e all'emergere di connessioni tra il cervelletto e la corteccia cerebrale. Pertanto, nei mammiferi, la parte ventrale del rombencefalo, il ponte (Varolii) (Fig. 175-15), comincia a sporgere, rappresentando una commissura di vie conduttrici, particolarmente significativa nell'uomo, che possiede la corteccia cerebrale più potente e il massima capacità di movimenti isolati degli arti. Il ponte si trova sotto forma di cresta trasversale davanti al midollo allungato. Con le sue estremità laterali si immerge nel cervelletto, formando le sue gambe laterali o laterali - brachia cerebelli lateralia.

A causa dell'omogeneità della funzione del cervelletto, la sua struttura istologica è uniforme (Fig. 162). Già in uno stato primitivo (nelle lamprede), il midollo grigio, che forma la corteccia cerebellare, è costituito da tre strati principali di cellule: cellule superficiali molecolari, granulari profonde e centrali di grandi dimensioni. Queste ultime nella serie degli animali sono altamente differenziate e sono conosciute come cellule di Purkinje con dendriti che si ramificano ad albero nello strato molecolare superficiale rigorosamente nel piano sagittale del corpo, mentre i dendriti delle cellule dello strato granulare si ramificano in quello segmentale aereo. Oltre alla corteccia, nel cervelletto dei mammiferi ci sono anche speciali gruppi di cellule che formano i nuclei cerebellari, e il più importante di essi è il nucleo dentato accoppiato - nucleo dentatus - che giace negli emisferi cerebellari (Fig. 163-18 ). C'è anche un numero di nuclei accoppiati situati tra quelli frastagliati. La corteccia cerebellare riceve impulsi dal midollo spinale e dal midollo allungato, dalle olive, dai nuclei uditivi del ponte e dalla corteccia cerebrale. I neuriti delle cellule corticali vengono inviati ai nuclei cerebellari (dentati e altri) e da qui al mesencefalo (nucleo rosso), al ponte e al midollo allungato. Attraverso questi centri, gli impulsi dal cervelletto vengono inviati attraverso il midollo spinale alla periferia.

Tutti i percorsi formano tre paia di peduncoli cerebellari: craniale al mesencefalo, laterale al ponte e caudale al midollo allungato, che corrisponde alla divisione del cervelletto in tre lobi principali: anteriore, medio e posteriore.
Mesencefalo
Il mesencefalo - mesencefalo (Fig. 159 e 160-3) - negli animali inferiori e nello stato embrionale negli animali superiori raggiunge dimensioni significative rispetto ad altre parti del cervello. Tuttavia, nelle forme adulte superiori è molto meno sviluppato. Solo nelle lamprede la volta mesencefalica è parzialmente rappresentata da una placca epiteliale, mentre nelle missine e in tutti gli altri vertebrati la volta - tectum opticum - è costituita da tessuto nervoso prodotto dalla placca laterale embrionale. Di conseguenza, la cavità cerebrale si riduce notevolmente, trasformandosi nelle forme superiori in un acquedotto canale-encefalico che collega il quarto ventricolo cerebrale del rombencefalo con il terzo ventricolo cerebrale del diencefalo. Il bordo anteriore del mesencefalo è un fascio di fibre nervose - la commissura posteriore - commissura posteriore. Negli animali inferiori, nel fornice sono presenti due tubercoli o lobi ottici, lobi ottici, che sono il centro visivo; in essi terminano le fibre del nervo ottico. Nei rettili, parte, e nei mammiferi, quasi tutte le fibre del nervo ottico terminano nel diencefalo, quindi i lobi ottici sono ridotti rispetto al talamo visivo (Fig. 164). Allo stesso tempo, anche negli anfibi, i lobi posteriori sono istologicamente associati all'organo dell'udito e dell'equilibrio. Questi lobi sono macroscopicamente più nettamente distinti in alcuni rettili, negli uccelli e soprattutto nei mammiferi, che non hanno un collicolo, ma un quadrigemina - corpora quadrigemina (Fig. 182-20, 21). In alcuni mammiferi dotati di buon udito, i collicoli posteriori sono più grandi di quelli anteriori (nelle balene e nei carnivori) (Fig. 188-6), ma solitamente i lobi ottici anteriori sono più grandi di quelli posteriori.
A causa della materia grigia della placca principale embrionale, situata ventralmente alla cavità del mesencefalo, si forma il cappuccio - tegmentum pedunculi (Fig. 187-3), costituito dai nuclei dei nervi oculomotore e trocleare - nuclei nervi III e IV - e il nucleo motore del cappuccio - nucleo motorius tegmenti. Quest'ultimo serve come continuazione di una massa cellulare simile del centro del midollo allungato, visibile nel cervello degli animali più primitivi. A partire dai rettili, è costituito da un numero di nuclei individuali. Il più importante di essi è il nucleo rosso, nucleo ruber (7); termina le fibre nervose che formano il peduncolo anteriore del cervelletto. L'altro nucleo interpeduncolare, il nucleo interpeduncolare, è collegato all'organo olfattivo attraverso il ganglio del frenulo.
Ventro-lateralmente tra il cervello e il romboide e il midollo spinale, alla base del mesencefalo nei mammiferi, c'è uno strato di percorsi - i peduncoli del cervello - redunculi cerebri (Fig. 175-16. 187-5). Sono più potenti nei mammiferi superiori. che è in relazione con lo sviluppo del mantello. Nelle parti mediali delle gambe ci sono le vie motorie e nelle parti laterali ci sono le vie sensoriali.

Il mesencefalo riceve impulsi dal cervelletto e dagli organi della vista, e negli animali terrestri, inoltre, dai cara degli emisferi cerebrali e dagli organi dell'udito e dell'equilibrio, invia esso stesso impulsi al midollo spinale e al midollo intermedio.
Diencefalo
Il diencefalo (Fig. 159 e 160-2) occupa un'area abbastanza significativa del cervello con un'ampia cavità del terzo ventricolo. Successivamente, però, la cavità ventricolare assume la forma di una fessura.
La placca tegmentale funge da volta per il terzo ventricolo, che in tutti gli animali rimane rudimentale, costituito da una placca epiteliale - lamina epithelialis - che, fusa (come nel cervello romboidale) con la pia madre, forma il tetto vascolare del terzo ventricolo. ventricolo cerebrale - tela chorioidea ventriculi tertii, che racchiude il plesso coroideo. Il tegmento viene introdotto mediante processi nella cavità del terzo ventricolo e, attraverso il foro interventricolare, penetra anche nel telencefalo, dove passa nei plessi coroidei dei ventricoli cerebrali laterali (Fig. 186-4, 10), formati a causa di la placca tegmentale del telencefalo.
I derivati della volta sono: 1) un'escrescenza a forma di tubo spaiata - l'epifisi e 2) un nodo accoppiato del frenulo.
L'epifisi, o ghiandola pineale, - l'epifisi (Fig. 159-6) è un rudimento del terzo, il cosiddetto occhio parietale. La ghiandola pineale, presente in quasi tutti gli animali, non è ugualmente sviluppata in tutti ed è assente solo in alcuni animali (marsupiali e alcuni altri).
Nei vertebrati inferiori, il diencefalo non svolge un ruolo come nell'amniota, motivo per cui in essi è relativamente poco sviluppato. Solo con lo spostamento dei centri visivi dal mesencefalo ad esso (Fig. 164), a causa del passaggio allo stile di vita terrestre, il diencefalo inizia ad aumentare, lasciando il mesencefalo molto indietro, cosa particolarmente evidente negli esseri umani. A causa della presenza di un numero significativo di nuclei di materia grigia, il diencefalo diventa un centro di correlazione per molte vie che vanno alla corteccia cerebrale e ritorno; È quindi chiaro che la differenziazione del diencefalo comincia dal momento della crescita del telencefalo.
Il diencefalo contiene una serie di centri vegetativi: termico, vasomotore, secretorio, metabolismo delle proteine e dei carboidrati, acqua, ecc.
Cervello finito
Il telencefalo (Fig. 159 e 160-1, 9) è una parte del cervello che in molti animali progredisce in modo estremamente forte e raggiunge il suo massimo sviluppo nella classe dei mammiferi, soprattutto nei primati. Negli esseri umani, il telencefalo supera così tanto tutte le altre parti del cervello da coprirle quasi tutte. Il telencefalo è diviso nel cervello olfattivo - rinencefalo - e nel pallio - pallium.
Cervello olfattivo
Dalla parete anteriore del telencefalo crescono un paio di sporgenze - bulbi olfattivi - bulbi olfactorii (Fig. 159-10), - che si collegano al cervello con una base ampia o gambe più o meno lunghe - tratti olfattivi - tractus olfactorius (in ossa pesci e rettili).
La base, o parte basale, del cervello assume dimensioni significative già nel cervello primitivo. Serve come centro olfattivo secondario ed è formato da un notevole ispessimento - il ganglio della base - nucleo basale (1), da cui nei mammiferi lo striato - corpo striato - sporge nella cavità dei ventricoli laterali del cervello. Il midollo grigio dello striato è diviso durante lo sviluppo dal midollo bianco, che forma la capsula interna - capsula interna (Fig. 184-9), - nella parte dorsale, o corpo caudato - nucleo caudatus, - e nella parte ventrale, o nucleo lentiforme (2) -nucleus lentiformis, e la capsula esterna -capsula externa (4) - separa la parte laterale, o recinzione (3), -claustrum. A causa della forte crescita, i corpi striatali, premendo nella cavità dei ventricoli laterali, li trasformano in spazi a fessura (5).
Nel cervello altamente differenziato dei mammiferi, la sua parte olfattiva è sviluppata in un numero di animali tutt'altro che uniforme.
Negli animali macrosmatici (compresi tutti gli animali domestici e, in generale, la maggior parte degli animali che hanno un senso dell'olfatto ben sviluppato), la parte olfattiva del cervello è piuttosto significativa.
Negli animali microsmatici (scimmie, odontoceti, delfini, capodogli), secondo il funzionamento dell'organo olfattivo, la parte olfattiva del cervello è piccola.
Delle singole parti del cervello olfattivo, i bulbi olfattivi sono particolarmente prominenti, che negli animali macrosmatici sono appendici del cervello sporgenti dalla parte anteriore (Fig. 177-I), e negli animali microsmatici sono molto piccoli. La parte esterna (ventrale) del ganglio basale, o corteccia olfattiva, forma davanti al chiasma ottico, tra il tratto olfattivo laterale e quello mediale, il tuberculum olfattivo olfattivo, che tra gli animali domestici è più pronunciato solo nei suini; in altri è piatto, per questo viene chiamato semplicemente triangolo olfattivo. Negli animali microsmatici l'area corrispondente sporge appena.
La struttura della corteccia cerebrale
Il mantello secondario è costituito da materia cerebrale grigia e bianca. Il primo si trova superficialmente e forma la corteccia cerebrale, il secondo si trova in profondità.
La corteccia cerebrale nelle sue varie parti è strutturata istologicamente in modo diverso, ciò che fu stabilito per primo da V. A. Betz (1874) e spiegò anche: "Questa differenziazione strutturale della corteccia cerebrale è un'espressione della localizzazione delle funzioni nella corteccia" (1881). . La differenza riguarda sia la localizzazione degli elementi cellulari (citoarchitettura) sia il decorso delle fibre nel midollo grigio (mieloarchitettura). La citoarchitettura e la mieloarchitettura della corteccia cerebrale sono state studiate più o meno dettagliatamente solo negli esseri umani e piuttosto scarsamente negli animali.
Gli elementi cellulari della corteccia si trovano parallelamente alla superficie del cervello in sei strati (strati). Questi strati, contando dall'esterno verso l'interno, sono i seguenti: I-molecolare, II-granulare esterno, III-strato di piccole cellule piramidali, IV-strato granulare interno, V-strato di grandi cellule piramidali e VI-strato di cellule polimorfiche ( Fig. 166). Nella serie filogenetica degli animali, non tutti e sei gli strati sono nettamente delimitati, ma la loro presenza può essere dimostrata in tutti i mammiferi.

Le cellule che formano la corteccia sono divise in tre tipi: Ramon-Kakhalevskij, piramidali e polimorfiche. Di queste, le cellule piramidali di Betz meritano un'attenzione speciale: più l'animale è organizzato o vecchio, più processi hanno, quindi le connessioni di queste cellule con altre sono più estese (Fig. 167).
A diversi strati cellulari vengono assegnate funzioni diverse: lo strato granulare interno (IV), che è quello primario, ha una funzione recettoriale, le cellule degli strati V e VI, le grandi cellule piramidali e fusiformi hanno una funzione effettrice. Successivamente, le cellule emergenti degli strati II e III svolgono funzioni associative di ordine superiore, cioè mentali.
Sulla base delle differenze locali di citoarchitettura, la corteccia dei mammiferi è divisa in regioni principali - campi - aree - e ogni campo, almeno nei mammiferi superiori e soprattutto nell'uomo, sulla base di diverse mieloarchitettoniche può essere ulteriormente suddiviso in un numero delle aree mieloarchitettoniche.
Filogeneticamente, tutti i campi del mantello secondario sono differenziati dalle quattro regioni primitive dei mammiferi inferiori (marsupiali e insettivori); queste aree sono diverse nella loro citoarchitettura e, a giudicare dalla presenza in esse di alcune fibre di proiezione (Fig. 165), corrispondono anche a centri funzionali speciali. Pertanto, nella regione frontale si trova il centro corticale motorio primitivo, nel pentro visivo occipitale, tra loro c'è il pentro sensoriale cutaneo e nella regione temporale c'è il pentro uditivo. I centri fisiologici e architettonici non sempre corrispondono alla posizione dei giri.
Il rapporto tra le fibre della polpa nella corteccia cerebrale è in una certa misura determinato dalla divisione accettata di esse in gruppi corrispondenti a sei strati di cellule (Fig. 168, a destra).

In base allo sviluppo della guaina mielinica (mielogeneticamente), tutte le fibre nervose sono classificate come sistema di proiezione o sistema di associazione. I primi si sviluppano prima della nascita, i secondi solo dopo la nascita. Mielogeneticamente la corteccia può essere divisa anche in base a due caratteristiche - la percentuale di fibre di proiezione e di associazione e la loro ontogenesi - in aree: primarie, terminali e intermedie, cioè fibre che sono rivestite dalla guaina mielinica prima della nascita, dopo la nascita e nella ultimo periodo dello stato embrionale.
Attualmente, sulla base dello studio della cito e mieloarchitettura della corteccia cerebrale, negli esseri umani vengono identificati più di 250 campi.
Le fibre di associazione collegano tra loro numerose aree della corteccia all'interno di un emisfero. Si dividono in brevi e lunghi; le prime collegano circonvoluzioni vicine, le seconde collegano aree e lobi più distanti tra loro. Le fibre lunghe sono particolarmente sviluppate nell'uomo, mentre negli animali sono insignificanti o completamente assenti.
Le fibre commissurali collegano aree situate in diversi emisferi. Formano le commissure anteriore e posteriore (commissure): la commissura del corno di Ammon e il corpo calloso.
Le fibre di proiezione sono divise anatomicamente in corte e lunghe e funzionalmente in corticofughe e cortico-petalo. Brevi fibre di proiezione collegano la corteccia con i nuclei del collicolo visivo e del tratto quadrigemino. Lunghi tratti collegano la corteccia con i nuclei pontini o il midollo spinale. Tutti questi percorsi, sia brevi che lunghi, passano tra il corpo caudato e i nuclei delle collinette ottiche, da un lato, e il nucleo lenticolare, dall'altro, formando la capsula interna - capsula interna (Fig. 184-9) . Nel mesencefalo queste fibre formano i peduncoli cerebrali.
Le vie di proiezione, così come quelle associative, sono più sviluppate negli esseri umani; Negli animali alcuni di essi sono addirittura completamente assenti.
Pertanto, gli emisferi cerebrali in generale e la corteccia cerebrale in particolare sono strutturati in modo più complesso e, inoltre, ineguale tra i mammiferi, raggiungendo il loro massimo sviluppo nei primati superiori e soprattutto nell'uomo. Secondo I.P. Pavlov, gli emisferi cerebrali sono continuamente bombardati da innumerevoli irritazioni sia dal mondo esterno che dall'ambiente interno del corpo stesso. Poiché la corteccia cerebrale degli animali superiori è portatrice della funzione di chiusura (cioè della funzione di acquisizione, della formazione di nuove connessioni tra l'organismo e l'ambiente, dello sviluppo di nuove esperienze di vita, della funzione di adattamento ontogenetico che adatta l'organismo alle condizioni ambientali), il dipartimento superiore del sistema nervoso centrale possiede la conoscenza di tutti i fenomeni che si verificano nel corpo.
Peso del cervello
Il peso assoluto del cervello degli animali varia notevolmente. È determinato dalle dimensioni dell'animale, dalla sua età, dalla sua specie, ecc. I cervelli più grandi si trovano negli animali di grandi dimensioni. Il suo peso raggiunge: balene - 4.673-7.000,0, elefanti - 4.370-5.430,0, cavalli - 372-570,0, bovini - 410-550,0, pecore - 97 -112,0, maiali - 96-145,0, cani - 46 -138,0, nei topi - 0,37. Negli esseri umani, il peso assoluto del cervello è molto elevato: 1.350-1.450,0.
Il peso relativo del cervello è generalmente inversamente proporzionale al peso dell'animale: nelle balene solo 1: 10.590 e anche 1: 14.000, negli elefanti - 1: 375-560, nei cavalli - 1: 480-1.000 (1), nei suini - 1: 1.200-1.900, nei cani - 1: 30-400. Negli esseri umani, il peso relativo del cervello è 1: 35-45. Negli animali giovani, il peso relativo del cervello è significativamente più alto che negli adulti; nei piccoli animali il peso del cervello è molto grande: nella scimmia scura e tenace è 1:15.
Il rapporto tra il peso del midollo spinale e del cervello: negli esseri umani - 1: 40, nei cavalli - 1: 2,5, nei tori - 1: 1,5, nei cani - 1: 4,5-9. Pertanto, il peso del midollo spinale negli esseri umani è solo il 2,5% del cervello e nei cavalli è del 40%.
C'è più materia grigia nei piccoli animali, ad esempio, la percentuale di materia grigia rispetto a quella bianca è 52,1 e 47,9 nei cavalli, 50,1 e 49,9 nei bovini, 61,1 e 38,9 nei cani, e il furetto addirittura 80,7 e 19,3 (Popov). .
- In contatto con 0
- Google+ 0
- OK 0
- Facebook 0






