"Antibiotici e chemioterapia", 2003, 48(10): 32-39.
Articolo pubblicato con il gentile permesso di Olga Efremenkova
Vladimirovna, testa settore ricerca composti naturali
Istituto di ricerca per la scoperta di nuovi antibiotici. G.F. Gause RAMS
Segnali comunicativi dei batteri
V.D. georgiano
Istituto di ricerca per la scoperta di nuovi antibiotici. G. F. Gause RAMS, Mosca
V.D. Georgia. Segnali comunicativi dei batteri
G.F. Istituto Gause di Nuovi Antibiotici, Accademia Russa delle Scienze Mediche, Mosca
Attualmente si assiste al passaggio dall’idea tradizionale dei batteri come organismi strettamente unicellulari all’idea delle comunità microbiche come strutture integrali che regolano le loro risposte comportamentali in base ai cambiamenti delle condizioni ambientali.
Colonie di quasi tutti i tipi di batteri dimostrano la capacità di differenziazione cellulare e di organizzazione multicellulare. Questa capacità è più evidente quando i batteri crescono nei loro habitat naturali, dove formano varie strutture multicellulari: biofilm, tappeti batterici, corpi fruttiferi, ecc.
Il concetto di Quorum Sensing è stato introdotto nel 1994. Significa la percezione da parte delle cellule dei cambiamenti nell'ambiente che si verificano quando una coltura batterica raggiunge un certo numero di soglia e la reazione a questi cambiamenti.
I processi descritti che si verificano solo con una densità di popolazione sufficientemente elevata includono i seguenti fenomeni:
- Bioluminescenza nei batteri marini Vibrio fisheri E V.harveyi;
- aggregazione delle cellule dei mixobatteri e successiva formazione di corpi fruttiferi con spore;
- sporulazione in bacilli e attinomiceti;
- stimolazione della crescita degli streptococchi e di numerosi altri microrganismi;
- coniugazione per trasferimento plasmidico Enterococcus faecalis e specie affini, nonché batteri del genere Agrobatterio;
- sintesi di esoenzimi e altri fattori di virulenza nei patogeni vegetali ( Erwinia carotovora, E.hyacinthii ecc.) e animali ( Pseudomonas aeruginosa, Staphylococcus aureus);
- formazione di antibiotici nei membri del genere Streptomiceti e a E. carotovora;
- formazione di biofilm nel R. aeruginosa e altri microrganismi.
Vengono divulgati i meccanismi di molti di questi processi, vengono determinati i fattori di comunicazione intercellulare responsabili dei processi che dipendono dalla densità di popolazione.
Un problema serio nella pratica clinica è la diffusa comparsa di forme resistenti di microrganismi, che riduce l'efficacia dell'uso di farmaci antibatterici. Di particolare difficoltà è l’aumento della resistenza ai farmaci dei batteri nei biofilm. I batteri spesso utilizzano reazioni di rilevamento del quorum per sintetizzare fattori di virulenza, antibiotici e formare biofilm. Pertanto, lo studio dei meccanismi di tali reazioni apre nuove opportunità per la prevenzione e il trattamento delle malattie causate da agenti microbici e consente anche di dare uno sguardo diverso al complesso complesso delle interazioni batteriche interspecifiche negli habitat naturali dei microrganismi.
I meccanismi delle reazioni di rilevamento del quorum differiscono nei batteri gram-positivi e gram-negativi, quindi è consigliabile considerarli separatamente.
Reazioni di rilevamento del quorum nei microrganismi Gram-positivi
I batteri Gram-positivi solitamente comunicano utilizzando molecole di segnalazione oligopeptidiche. La segnalazione nella maggior parte dei casi coinvolge un meccanismo di fosforilazione a due componenti. Di norma, lo stato di quorum viene raggiunto quando la popolazione di cellule batteriche entra nella fase stazionaria di crescita. È in questo momento che vengono rilevate le molecole segnale, con l'aiuto delle quali le cellule entrano in contatto tra loro. Lo schema generale di comunicazione dei batteri gram-positivi può essere rappresentato come segue: in primo luogo, nella cellula viene sintetizzato un precursore che, essendo modificato, si trasforma in un oligopeptide maturo. Quest'ultimo viene escreto all'esterno della cellula dall'esportatore. Le molecole di oligopeptidi si accumulano nello spazio intercellulare man mano che aumenta la densità delle cellule batteriche. Una chinasi sensoriale bicomponente che penetra nella membrana riconosce il segnale e lo trasmette nella cellula attraverso una cascata di fosforilazione. Nella cellula, la molecola oligopeptidica interagisce con il gene(i) bersaglio.
Il classico sistema quorum-dipendente del peptide può essere considerato il sistema responsabile del trasferimento coniugativo dei plasmidi in Enterococcus faecalis e specie batteriche affini. Questo sistema stimola la distribuzione nella popolazione microbica di tratti importanti per l'interazione tra il microrganismo e l'animale ospite, nonché per l'eliminazione della competizione. Il plasmide pPDl trasportato dal sistema quorum-dipendente del peptide è responsabile della sintesi delle emolisine, il plasmide pCDl è responsabile della formazione della batteriocina, il plasmide pCFlO è responsabile della resistenza E.faecalis alla tetraciclina. Ciascun esa- o octa-peptide induce l'adesione delle cellule batteriche e la loro coniugazione con il trasferimento dal donatore al ricevente di uno specifico plasmide. Ad esempio, l'ottapeptide cPDl stimola il trasferimento coniugativo del plasmide pPDl. Il plasmide codifica per un recettore situato sulla proteina repressore dell'operone corrispondente. L'interazione dell'oligopeptide con il recettore provoca la dissociazione del repressore dal DNA, innescando così la sintesi del prodotto corrispondente. Il plasmide pPDl comprende anche il gene traC, il cui prodotto è una proteina che facilita la penetrazione del peptide attraverso la parete cellulare. I segnali oligopeptidici vengono sintetizzati intensamente da cellule che non portano i corrispondenti plasmidi (riceventi), mentre la sintesi di tali segnali è soppressa nelle cellule donatrici; inoltre, il plasmide codifica per un peptide inibitorio.
Il prodotto del plasmide pPDl è il peptide iPDl, HHaKTHBHpyiounmcPDl.
Un altro processo dipendente dal quorum trovato in E.faecalis, è la produzione di due fattori di virulenza: gelatinasi (GelE) e serina proteasi (SprE).
Un esempio dell'uso di un segnale peptidico per le interazioni intercellulari è il sistema di rilevamento del quorum, che controlla la sintesi delle esotossine nella fase logaritmica tardiva della crescita in Staphylococcus aureus. In questo sistema, la proteina AgrD viene sintetizzata come un precursore costituito da 46 aminoacidi, che, durante l'esportazione da parte della proteina AgrB, viene convertito in un AIP maturo (peptide autoinducente) composto da 8 aminoacidi. L'AIP è riconosciuto dal sensore chinasi a due componenti AgrC, che trasmette un segnale nella cellula durante la fosforilazione del regolatore di risposta, AgrA. AgrA~P attiva la trascrizione dei geni bersaglio, stimola la trascrizione dell'operone agrB, D, C, A (circuito autoregolatorio positivo) e “proibisce” anche la trascrizione dei geni che codificano per altre esotossine. Basato sulle differenze nell'AIP e nel suo recettore, i ceppi S. aureus può essere assegnato a quattro o più gruppi. Gli oligopeptidi sintetizzati da uno dei gruppi inducono patogenicità in questo gruppo e sopprimono specificamente i sistemi di virulenza Agr in altri gruppi.
L’emergere della competenza nella fase logaritmica tardiva della crescita dipende dalla densità della popolazione. Streptococco pneumoniae. Il gene comC codifica per un precursore costituito da 41 residui aminoacidici. Quest'ultimo viene convertito in un peptide maturo costituito da 17 residui di aminoacidi nel processo di interazione con il sistema di esportazione del peptide (sistema ABC), formato dai prodotti dei geni comAB. Il peptide contatta il suo recettore sulla superficie cellulare, l'istidina chinasi, un prodotto del gene comD. L'istidina chinasi attivata fosforila il prodotto del gene comE. Man mano che le cellule si accumulano, il numero di segnali peptidici aumenta e raggiunge un livello critico nel mezzo. Di conseguenza aumenta anche la quantità di proteina comE fosforilata che, a partire da una certa concentrazione, si lega al promotore dell'operone comCDE, stimolandone il lavoro (circuito autoregolatorio positivo), attiva il promotore dell'operone comAB (il sistema di esportazione delle proteine dalla cellula) , attiva l'operone comX, che comprende l'intera catena dei geni della competenza tardiva; responsabile del legame e dell'assorbimento del DNA in trasformazione e di tutti gli altri stadi tardivi della trasformazione.
Oltre agli esempi sopra riportati di reazioni di rilevamento del quorum nei batteri gram-positivi, va notato che, oltre agli oligopeptidi, i batteri gram-positivi utilizzano anche sostanze di diversa natura chimica come molecole di segnalazione. Quindi, i rappresentanti dell'ordine Actinomiceti Insieme alle molecole segnale peptidiche, sono state trovate sostanze di natura a basso peso molecolare, la maggior parte delle quali contiene un gruppo lattone.
Negli streptomiceti, i sistemi di rilevamento del quorum coinvolgono i butirrolattoni e i loro corrispondenti recettori proteici, che insieme regolano lo sviluppo morfologico e la produzione di antibiotici nei loro produttori. Il regolatore degli actinomiceti più studiato è il fattore A, che è il 2-iso-capriloil-3-idrossimetil-y-butirrolattone.
L'influenza del fattore A sulla differenziazione morfologica e sulla formazione di antibiotici è soggetta allo schema generale di funzionamento dei regolatori degli streptomiceti contenenti un gruppo lattone. Nelle prime fasi della crescita, quando la concentrazione del fattore A è bassa, il recettore del fattore A (AgrA) si lega e reprime l'espressione dell'ipotetico attivatore comune della biosintesi e della sporulazione della streptomicina. Isolamento di AgrA dal lisato cellulare S. griseusÈ stato dimostrato che l'IFO 13350 è una proteina di 276 aminoacidi con un peso molecolare di 29,1 kDa.
All’aumentare della densità della coltura, la concentrazione del fattore A raggiunge un livello critico al quale si lega ad AgrA, provocando la dissociazione di quest’ultimo dal DNA e attivando così la trascrizione del gene chiave adpA che codifica per AdpA (una proteina di 405 aminoacidi contenente un legame sito nella regione centrale) con DNA, simili ai regolatori della trascrizione della famiglia delle proteine AraC/XylS). AdpA, a sua volta, è un regolatore positivo dell'attivatore citoplasmatico rilevato del cluster genetico della biosintesi della streptomicina e degli attivatori del processo di sporulazione. L'attivatore citoplasmatico, legandosi al DNA nella regione del promotore del gene per la regolazione specifica del cluster di biosintesi della streptomicina strR, induce la trascrizione di questo gene, il gene per la resistenza al proprio antibiotico - aphD, situato dopo di esso, l'adsA gene, che codifica per il fattore a extracitoplasmatico della RNA polimerasi, necessario per la formazione del micelio aereo, nonché per il gene sgmA che codifica per la proteina peptidasi, che, insieme ad altri enzimi idrolitici, è coinvolta nella degradazione del micelio substrato proteine come risultato della formazione del micelio aereo. Il prodotto regolatore del gene strR determina l'inizio della trascrizione dei geni della biosintesi strutturale come parte di un cluster di promotori StrR-dipendenti. L'inizio dell'espressione del promotore del gene strR sotto l'influenza di un attivatore citoplasmatico garantisce anche la produzione del prodotto del gene aphD, l'aminoglicoside fosfotransferasi, e quindi la creazione di un livello base di resistenza del ceppo al proprio antibiotico.
È stato dimostrato che in diverse specie di streptomiceti esiste omologia tra gli elementi strutturali dei regolatori. Sequenze nucleotidiche omologhe al gene agrA in S. griseus, si trovano anche in altri streptomiceti. Ad esempio, a S.coelicolor A3 (2), sono stati trovati due geni srA e srB, che codificano per le proteine simili ad AgrA CrA e CrB, che sono simili al 90,7% tra loro e al 35% ad AgrA.
Reazioni di rilevamento del quorum nei microrganismi Gram-negativi
In più di 450 specie di batteri gram-negativi sono stati trovati sistemi quorum-dipendenti, in cui vari lattoni acilhomoserina fungono da molecole di segnalazione. Lo schema generale di comunicazione nei batteri gram-negativi può essere rappresentato come segue: nel sistema di rilevamento del quorum dei batteri gram-negativi, le proteine della famiglia Luxl sono sintasi autoinduttrici e catalizzano la formazione di specifiche molecole autoinduttrici di acil omoserina lattone. Gli autoinduttori si diffondono liberamente attraverso la membrana e si accumulano all’aumentare della densità cellulare. Le proteine della famiglia LuxR si legano ai relativi autoinduttori quando viene raggiunta una concentrazione sufficientemente elevata di molecole di segnalazione. Il complesso LuxR, un autoinduttore, si lega al promotore dei geni bersaglio, iniziandone la trascrizione.
Batteri del genere Erwinia (E. carotovora, E. chrysanthemii) sono agenti patogeni delle piante. Abbattono le pareti cellulari delle piante con l'aiuto di pectinasi e cellulasi. La formazione di questi enzimi è un importante fattore di virulenza e dipende dalla densità della popolazione. A Erwinia il sistema genetico expI-expR funziona in modo simile al sistema luxI-luxR in V.fisheri. Le reazioni di quorum sensing coinvolgono anche il sistema regolatorio fornito dalla trascrizione dei geni rsmA-rsmB. Dalla densità di popolazione E. carotovora dipende anche dalla sintesi dell'antibiotico carbapenemico. La produzione di questo antibiotico è sotto il controllo del cluster di geni carA-carH e potrebbe essere necessaria per eliminare i microrganismi concorrenti nel locus dell'infezione della pianta.
Viene mostrato un altro esempio dell'uso dei lattoni omoserina come molecole di segnalazione Pseudomonas aeruginosa- un agente patogeno per gli animali. Patogenicità dentro R. aeruginosa a causa di un ampio arsenale di fattori di virulenza. Alcuni di essi sono associati alla cellula (pili, adesine, lipopolisaccaridi), altri vengono secreti (proteasi, ramnolipidi, esoenzima S, esotossina A, antibiotico piocianina, ecc.). La formazione di molti dei fattori di virulenza extracellulare è controllata da sistemi di interazione intercellulare. I componenti centrali di tali interazioni sono i sistemi di rilevamento del quorum las e rhl, che attivano l'espressione genica a seconda della densità cellulare del microrganismo. Ciascun sistema è rappresentato da due geni: uno codifica per un enzima che sintetizza uno specifico autoinduttore: l'omoserina lattone acilato (lasl/rhll); l'altro codifica per un attivatore di trascrizione al quale si lega il corrispondente autoinduttore (lasR/rhIR). L'autoinduttore dei sistemi las e rhi è l'N-(3-ossododecanoil)-L-omoserina lattone (3-oxo-C12-HSL), che viene esportato dalla cellula da un sistema speciale chiamato MexEF-OprN-pump e N- butirril-L-omoserina lattone (C4-HSL), rispettivamente.
Il sistema LAS controlla l'espressione dei geni che codificano per fattori di virulenza come l'elastasi A, B e la proteasi alcalina; sistema rhi - enzimi per la biosintesi dei ramnolipidi, piocianina. Recentemente è stata scoperta una terza molecola di segnalazione coinvolta nelle reazioni di rilevamento del quorum P.aeruginosa- 2-eptil-3-idrossi-4-chinolone (PQS). Questa molecola di segnalazione può controllare il livello di espressione di las B, che codifica per l'elastasi Las B, nonché il livello di espressione di rhil, che codifica per C4-HSL sintasi.
Batterio Agrobacterium tumefaciens provoca la formazione di galle della corona in molte specie vegetali. Le galle sono un analogo vegetale di un tumore maligno e si formano a seguito del trasferimento di frammenti di DNA oncogenico da un batterio nel nucleo di una cellula vegetale mediante plasmidi Ti. Alcuni dei geni del plasmide Ti determinano la sintesi delle opine da parte delle cellule vegetali, che fungono da substrato nutritivo per A.tumefaciens. Il sistema genetico traI-traR omologo luxI-luxR stimola la diffusione dei plasmidi Ti nella popolazione batterica. Il DNA plasmidico tende a diffondersi in tutta la popolazione batterica e, una volta creato un "quorum" sufficiente, induce le cellule portatrici di plasmide a coniugarsi con altre cellule batteriche. Allo stesso tempo, il trasferimento coniugativo dei plasmidi Ti dipende dalle opine. In particolare, la trascrizione di traR è stimolata dal fattore OccR attivato dall'octopina.
Rilevamento del quorum nelle formazioni multicellulari
La capacità dei batteri di formare biofilm è interessante in considerazione del fatto che i rappresentanti di agenti patogeni per l'uomo e gli animali mostrano resistenza all'azione delle sostanze antimicrobiche quando crescono nei biofilm. I biofilm sono comunità batteriche altamente ordinate che consentono ai batteri di vivere in uno stato attaccato. I biofilm possono essere composti da uno o più tipi di batteri. Sono permeati da una rete di canali d'acqua che assicurano la fornitura di nutrienti ai membri della comunità e rimuovono i prodotti metabolici. All'interno di un singolo biofilm si possono osservare diversi modelli di espressione genetica, suggerendo che i singoli membri della comunità hanno "responsabilità specifiche" che, se combinate con altre, migliorano la vitalità dell'intero consorzio.
I biofilm si formano nei polmoni da un agente patogeno P.aeruginosa. Lo spessore di un tale biofilm è di diverse centinaia di micrometri. Le microcolonie in un biofilm maturo si trovano nella matrice polisaccaridica extracellulare. All'interno del biofilm si trova eterogeneità: in esso c'è un gradiente di ossigeno - una diminuzione della concentrazione di ossigeno dalla periferia verso l'interno. Si prevede che gradienti simili siano riscontrabili per il pH e i nutrienti. Questi gradienti forniscono variabilità fisiologica tra le singole cellule del biofilm: ad esempio, le cellule crescono molto più lentamente in profondità che alla periferia. Il batterio in un biofilm così maturo è fenotipicamente resistente agli agenti battericidi. Pertanto, i biofilm causano vari tipi di infezioni batteriche croniche. Formazione di biofilm nel P.aeruginosaè sotto il controllo delle reazioni di rilevamento del quorum. Le mutazioni nel gene lasI compromettono la maturazione del biofilm, poiché la proteina LasI non sintetizza 3-oxo-C12-HSL e la formazione del microfilm non continua dopo la fase di microcolonia. Il ruolo di C4-HSL nei processi di formazione rimane sconosciuto. I biofilm formati dai mutanti della proteina LasI sono sensibili ai detergenti, mentre i biofilm normali sono resistenti. Ciò dà motivo di pensare che la terapia mirasse alla disregolazione del meccanismo di rilevamento del quorum P.aeruginosa, può portare all'arresto della formazione del biofilm, che aumenterà la sensibilità di questo batterio agli agenti antimicrobici.
Formazione di un biofilm in un batterio patogeno Burkholderia cepacia definito anche dal “senso del quorum”. Quando cresce nei biofilm, questo microrganismo è simile a P.aeruginosa mostra una significativa resistenza agli agenti antimicrobici.
Interazioni interspecifiche dei microrganismi
Le comunicazioni interspecifiche nei batteri possono servire a sincronizzare le funzioni specializzate delle specie in un gruppo. La diversità presente in ogni popolazione può migliorare la sopravvivenza dell’intera comunità. Inoltre, le interazioni produttive basate sul quorum sensing possono promuovere lo sviluppo di organizzazioni batteriche multispecie, come i biofilm, nonché la creazione di specifiche associazioni simbiotiche con ospiti eucariotici.
Le interazioni interspecifiche dei microrganismi sono state studiate in modo più approfondito utilizzando l'esempio della comunità microbica della cavità orale e della superficie dei denti umani. Sono state identificate circa 500 specie di batteri nei biofilm sulla superficie dei denti, che funzionano come una comunità coordinata con comunicazioni intra e interspecifiche. Gli streptococchi costituiscono dal 60 al 90% dei batteri che colonizzano la superficie dei denti durante le prime quattro ore dopo la pulizia dal dentista. Tra gli altri tipi di "primi colonizzatori" si trovano rappresentanti Actinomiceti, Capnocitofagi, Eikenella, Emofilo, Prevotella, Propionibatterio E Veillonella.
È probabile che le modalità di comunicazione tra cellule geneticamente identiche differiscano dai segnali nella comunicazione interspecie. Non ci sono prove della presenza di rappresentanti tipici della famiglia dei lattoni acilhomoserina tra le molecole di segnalazione dei batteri orali, che regolano l'espressione genica intraspecifica nei batteri Gram-negativi.
AI-2 è la principale molecola di segnalazione nelle comunicazioni interspecifiche. Ciò è confermato dalla scoperta del gene luxS che codifica l'enzima necessario per la sintesi della molecola AI-2 in diversi generi di batteri orali.
L'AI-2 è stato scoperto per la prima volta in un batterio luminoso marino Vibrio Harveyi, per la quale è una molecola segnale che regola il processo di bioluminescenza. Successivamente, la presenza di AI-2 è stata dimostrata in più di 30 specie di batteri, inclusi microrganismi gram-positivi e gram-negativi.
A volte può essere utile per un gruppo di batteri influenzare negativamente il ciclo di rilevamento del quorum di un gruppo di batteri concorrenti. La ricerca in questo settore rivela diversi esempi di strategie di rilevamento dell'antiquorum che utilizzano popolazioni batteriche coesistenti. COSÌ, Staphylococcus epidermidis utilizza il peptide per controllare il livello della sua virulenza agr, nonché per sopprimere la virulenza Staphylococcus aureus.
Sottoporre a tensione bacillo sp. 240B1 dimostra la capacità di inattivare enzimaticamente i lattoni acilhomoserina, le molecole di segnalazione dei batteri Gram-negativi. È stato dimostrato che in presenza dell'AIA, omoserina lattonasi, costituita da 250 aminoacidi, le molecole di omoserina lattoni prodotte dall'agente patogeno nelle piante vengono distrutte. Erwinia carotovora. Geni omologhi al gene aiiA sono stati trovati anche in 16 sottospecie Bacillus thuringiensis pertanto questi microrganismi sono in grado di degradare anche i lattoni dell'omoserina.
batterio del suolo Paradosso di Variovorax può utilizzare lattoni acilomoserina come unica fonte di carbonio e azoto. Questo fatto indica che nei loro habitat naturali V.paradosso possono crescere sui lattoni acilhomoserina, beneficiando dell’esacerbazione della concorrenza nell’ambiente. In questo caso, l'enzima che distrugge i lattoni acilomoserina è diverso dall'AiiA-lattonasi: si tratta di un'amminoacilasi che separa l'anello lattonico dal gruppo acilico.
Dato che i sistemi di rilevamento del quorum controllano la virulenza in molti agenti patogeni animali e vegetali, questi sistemi possono essere considerati potenziali bersagli per l’azione degli agenti antimicrobici. Innanzitutto, una strategia consiste nell'inibire la sintesi delle molecole precursori dell'acilhomoserina lattone o degli stessi lattoni acilhomoserina. In secondo luogo, i sistemi che controllano il rilascio e la diffusione degli acil-omoserina lattoni possono fungere da bersagli per i farmaci. In terzo luogo, gli antagonisti simili al lattone dell'acilomoserina possono competere con gli lattoni dell'acilomoserina per il legame con gli omologhi LuxR. In quarto luogo, è possibile utilizzare enzimi che scindono gli acil omoserina lattoni, nonché anticorpi contro queste molecole. E, infine, come è stato recentemente dimostrato, i geni aiiA che codificano per lattonasi che degradano i lattoni acilomoserina possono essere introdotti nel genoma della pianta, esprimendosi in cui potrebbero proteggere la pianta ospite dai microrganismi patogeni. Pertanto, le piante di tabacco transgeniche con un gene aiiA incluso hanno resistito con successo all'infezione E. carotovora.
Citochine batteriche
È stato scoperto che i microrganismi procarioti sintetizzano sostanze simili agli ormoni dei vertebrati (compresi gli steroidi e gli ormoni polipeptidici come l'insulina). Un numero crescente di prove evidenzia l’importanza delle interazioni cellula-cellula mediate chimicamente nelle colture batteriche per eventi quali sporulazione, coniugazione, virulenza e bioluminescenza. Pertanto, attualmente, molti studi nel campo della microbiologia sono dedicati alle interazioni tra microrganismi basate sull'uso di citochine batteriche.
È noto che i microrganismi sono in grado di adattarsi in modo flessibile alle mutevoli condizioni ambientali (in particolare alla mancanza di nutrienti). Allo stesso tempo, alcuni di loro hanno un'organizzazione specifica del metabolismo geneticamente fissata, che consente loro di esistere a concentrazioni molto basse di nutrienti (oligotrofi). Le cellule di un'altra categoria (copiotrofi), quando l'ambiente è esaurito, sono in grado di attivare programmi speciali per sperimentare condizioni avverse. Alcuni di essi formano strutture specializzate (spore e cisti) estremamente resistenti a vari stress, mentre i batteri non sporulanti sono in grado di sopravvivere a condizioni avverse, rimanendo cellule vegetative con attività metabolica ridotta, cioè. passando in uno stato speciale VBNC (vitale ma non coltivabile - vitale, ma non coltivato). Naturalmente, i batteri non coltivati rimangono fuori dall'ambito dei metodi di ricerca generalmente accettati (la semina su terreni solidi o liquidi non ne consente il rilevamento). Ad esempio, gli agenti causali di malattie pericolose come il colera e la campilobatteriosi tendono a formare forme incolte. L'esame microscopico di campioni isolati dall'ambiente (suolo, acque fluviali e marine, ecc.) ha rivelato molte cellule che, avendo attività metabolica, non possono formare una coltura completa (cioè incolte). Attualmente sono noti solo pochi esempi di trasformazione di tali batteri in normali cellule in coltura. Il concetto di crescita dei microrganismi citochina-dipendente ci consente di riconsiderare il problema della selezione dei terreni per il ripristino di forme non coltivabili.
Forme incolte di batteri patogeni si trovano non solo nell'ambiente, ma anche nei tessuti, negli organi dell'uomo e degli animali. Molto spesso sono molto diversi morfologicamente e biochimicamente. Ad esempio, l'agente eziologico della tubercolosi nei tessuti forma forme coccoidi atipiche. È possibile che tali cellule siano forme speciali sopravvissute in grado di attivarsi e riprodursi. L'esistenza di tali forme dormienti può spiegare le ricadute ricorrenti della malattia in pazienti apparentemente guariti. È stato dimostrato che le cellule Mycobacterium tuberculosis può trasformarsi in uno stato coccoide non replicante in condizioni microaerofile in vitro che spesso si verificano in vivo(ad esempio, nei granulomi). Sono state trovate anche forme coccoidi Campylobacter jejuni E Helicobacter pylori. Si presume che si formino nei tessuti in risposta agli effetti dei farmaci e, possibilmente, siano cellule quiescenti resistenti agli antibiotici. Tuttavia, i dati sulla coltivazione di tali forme sono molto contraddittori. È possibile che tali batteri possano essere attivati da alcuni specifici fattori di crescita, il cui ruolo è probabilmente svolto dalle citochine dell'ospite. Ad esempio, la crescita dei bacilli della tubercolosi all’interno dei monociti è stata significativamente stimolata dal fattore di crescita trasformante (TGF-1), mentre la crescita cellulare M.tubercolosi E M.avium all'interno dei macrofagi è stata significativamente accelerata in presenza del fattore di crescita epidermico. Ovviamente, i fattori citochinici dell'ospite possono svolgere un ruolo importante sia nell'attivazione di batteri dormienti che nella riproduzione di patogeni attivi. Una diminuzione del livello di insulina nel sangue dei pazienti con diabete mellito porta ad una significativa proliferazione delle cellule. Pseudomonas pseudomallei, che sono gli agenti causali della melioidosi, e la transferrina è di grande importanza per la crescita e la sopravvivenza delle cellule all'interno dei macrofagi del topo Francisella tularensis.
È possibile che anche specifiche citochine batteriche svolgano un ruolo significativo nella formazione di forme quiescenti e nel loro recupero in cellule attive in divisione. Quindi, tenendo conto dei problemi legati all'emergere della resistenza agli antibiotici, è difficile sopravvalutare l'importanza di trovare fattori di crescita autocrini necessari per la crescita di batteri patogeni e, quindi, di essere un bersaglio per l'azione di antibiotici fondamentalmente nuovi che sono non tossico per il paziente.
L'uso di citochine batteriche specifiche può anche migliorare significativamente la situazione con la coltivazione di batteri non coltivabili in terreni non del tutto adatti alla loro riproduzione. Ad esempio, i micrococchi che di solito non crescono su un mezzo succinato minimo iniziano a moltiplicarsi normalmente in presenza del fattore autocrino Rpf (fattore di promozione della rianimazione), mentre le cellule lavate Mycobacterium smegmatis, che crescono su terreno minimo solo aggiungendo Rpf estratto da Micrococco luteo, può essere considerato come un modello di una popolazione di batteri "affamati" nel suolo, che probabilmente richiede la presenza di una citochina specifica per iniziare la divisione. L'uso di citochine batteriche specifiche può anche migliorare significativamente la situazione con la coltivazione di batteri non coltivabili in terreni non del tutto adatti alla loro riproduzione. Geni simili al gene che codifica per la proteina Rpf M.luteus, sono ampiamente distribuiti tra i batteri Gram-positivi con un alto contenuto di G+C, che comprendono streptomiceti, corinebatteri e micobatteri. Questo fatto apre nuove possibilità per la prevenzione e il trattamento delle malattie causate da agenti microbici e ci consente anche di dare uno sguardo diverso al complesso complesso delle interazioni batteriche interspecifiche negli habitat naturali dei microrganismi.
Ultimo aggiornamento: 20/02/2004
LETTERATURA
- Oleskin A.V., Botvinko I.V., Tsavkelova E.A. Organizzazione coloniale e comunicazione intercellulare nei microrganismi. Microbiologia 2000; 69:3:309-327.
- Fuqua W.C., Winans S., Greenberg E. Quorum sensing nei batteri: la famiglia Lux R-Lux I di regolatori trascrizionali sensibili alla densità cellulare. J Bacteriol 1994; 176:2:269-275.
- Meighen E. Biologia molecolare della bioluminescenza batterica. Microbiol Rev 1991; 55:1:123-142.
- Winans SC, Bassler B.L. Psicologia mafiosa. J Bacteriol 2002; 184:4:873-883.
- Writh R., Muscholl A., Wanner G. Il ruolo dei feromoni nelle interazioni batteriche. Tendenze Microbiol 1996; 4:3:96-103.
- Khokhlov A.S. Autoregolatori microbici a basso peso molecolare. M.: 1988; 270.
- Waldburger C., Gonzalez D., Chambliss G.H. Caratterizzazione di un nuovo fattore di sporulazione in Bacillus subtilis. J Bacteriol 1993; 175: 6321-6327.
- Pestova E., Havarstein L., Morrison D. Regolazione della competenza per la trasformazione genetica in Streptococco pneumoniae da un feromone peptidico autoindotto e da un sistema regolatorio a due componenti. Mol Microbiol 1996; 21:4: 853-862.
- Alloing G., Martin B., Granadel G., Claveris J. Sviluppo delle competenze in Streptococco pneumoniae: autoinduzione dei feromoni e controllo del quorum-sensing da parte della permeasi oligopeptidica. Ibid 1998; 9:1:75-83.
- Prozorov A.A. Ferormoni di competenza nei batteri. Microbiologia 2001; 70:1:5-14.
- Salmond G., Bycroft B., Stewart C., Williams P. L'"enigma" batterico: decifrare il codice della comunicazione cellula-cellula. Mol Microbiol 1995; 16:4:615-624.
- Greenberg E., Winans S., Fuqua C. Rilevamento del quorum da parte dei batteri. Ann Rev Microbiol 1996; 50:727-751.
- Otto M., Sussmuth R., Vuong C. et al. Inibizione dell'espressione del fattore di virulenza in Staphylococcus aureus dal Staphylococcus epidermidis feromone agr e derivati. FEBBRAIO Lett. 1999; 450:257-262.
- Dong Y., Xu J., Li X., Zhang L. AiiA, un enzima che inattiva il segnale di rilevamento del quorum dell'acil-omoserina lattone e attenua la virulenza di Erwinia carotovoru. Proc Natl Acad Sci 2000; 97:7: 3526-3531.
- Byers J., Lucas C., Salmond G., Welch M. Fatturato non enzimatico di un Erwinia carotovora molecola di segnalazione quorum-sensing. J Bacteriol 2002; 184:4: 1163-1171.
- Calfee M., Coleman J., Pesci E. L'interferenza con la sintesi del segnale dei chinoloni di Pseudomonas inibisce l'espressione del fattore di virulenza mediante Pseudomonas aeruginosa. Proc Natl Acad Sci 2001; 98:20:11633-11637.
- Nakayama J., Takanami Y., Horii T. et al. Meccanismo molecolare della segnalazione dei feromoni specifici della marea peptidica Enterococcus faecalis: funzioni del recettore dei feromoni TraA e della proteina legante i feromoni TraC codificata dal plasmide pPDI. J Bacteriol 1998:180:3:449-456.
- Mylonakis E., Engelbert M., Qin X. et al. IL Enterococcus faecalis Il gene fsrB, un componente chiave del sistema di rilevamento del quorum fsr, è associato alla virulenza nel modello di endoftalmite del coniglio. Infetta Immun 2002; 70:8:4678-4681.
- Sifri C., Mylonakis E., Singh V. et al. Effetto di virulenza di Enterococcus faecalis geni della proteasi e locusta sensibile al quorum in Caenorhabditis elegans e topi. ibid 2002; 70:10: 5647-5650.
- Matson M., Armitage J., Hoch J., Macnab R. Locomozione batterica e trasduzione del segnale. J Bacteriol 1998; 180:5:1009-1022.
- Onaka H., Horinouchi S. Attività di legame al DNA della proteina del recettore del fattore A e sue sequenze di DNA di riconoscimento. Mol Microbiol 1997; 24:991-1000.
- Onaka H., Ando N., Nihira T., Yamada Y. et al. Clonazione e caratterizzazione del gene del recettore del fattore A da Streptomyces griseus. J Bacteriol 1995; 177:21:6083-6092.
- Ohnishi Y., Kameyama S., Onaka H., Horinouchi S. La cascata regolatoria del fattore A che porta alla biosintesi della streptomicina in Streptomyces griseus: identificazione di un gene bersaglio del recettore del fattore A. Mol Microbiol 1999; 34:102-111.
- Yamazaki H., Ohnishi Y., Horinouchi S. Un fattore sigma con funzione extracitoplasmatica dipendente dal fattore A (OAdsA) che è essenziale per lo sviluppo morfologico in Streptomyces griseus. J Bacteriol 2000; 182:16:4596-4605.
- Kato J., Suzuki A., Yamazaki H. et al. Controllo da parte del fattore A di un gene della metalloendopeptidasi coinvolto nella formazione del micelio aereo Streptomyces griseus. ibid 2002; 184:21:6016-6025.
- Onaka H., Nikagawa T., Horinouchi S. Coinvolgimento di due omologhi del recettore del fattore A in Streptomyces coelicolor A3(2) nella regolazione del metabolismo secondario e della morfogenesi. Mol. microbiolo. 1998; 28:4:743-753.
- Revenchon S., Bouillant M., Salmond G., Nasser W. Integrazione del sistema di rilevamento del quorum nelle reti regolatorie che controllano la sintesi dei fattori di virulenza in Erwinia chrysanthemii. Mol Microbiol 1998; 29:1407-1418.
- Chatterjee A., Cui Y., Chatterjee A.K. RsmA e il segnale di rilevamento del quorum, N--L-omoserina lattone, controllano i livelli di RNA rsmB nell'RNA Erwinia carotovora sost. carotovora compromettendone la stabilità. J Bacteriol 2002; 184:15: 4089-4095.
- Kohler T., Van Delden C., Curty L. et al. La sovraespressione del sistema di efflusso multifarmaco MexEF-OprN influenza la segnalazione cellula-cellula in Pseudomonas aeruginosa. Ibid 2001; 183:18:5213-5222.
- Gallagher L., McKnight S., Kuznetsova M. el al. Funzioni richieste per la segnalazione extracellulare dei chinoloni Pseudomonas aeruginosa. ibid 2002; 184:23:6472-6480.
- Parsek M., Greenberg P. Rilevamento del quorum del lattone acil-omoserina nei batteri gram-negativi: un meccanismo di segnalazione coinvolto nelle associazioni con organismi superiori. Proc Natl Acad Sci 2000; 97:16:8789-8793.
- Conway V.A., Wepi V., Speert D. Formazione di biofilm e produzione di acil-omoserina lattone nel Burkholderia cepacia complesso. J Bacteriol 2002; 184:20:5678-5685.
- Kolenbrander P., Andersen R., Blehert D. et al. Comunicazione tra batteri orali. Microbo. Biologia Molecolare Rev 2002; 66:3:486-505.
- Miller M., Bassler B. Quorum che rileva nei batteri. Annu Rev Microbiol 2001; 55:165-199.
- Frias J., Olle E., Alsina M. Gli agenti patogeni parodontali producono molecole di segnale di rilevamento del quorum. Infettare Immun 2001; 69:3431-3434.
- McNab R., Ford S., EI-Sabaeny A. et al. Segnalazione basata su LuxS in Streptococco gordonii: L'autoinduttore 2 controlla il metabolismo dei carboidrati e la formazione del biofilm Porphyromonas gingivalis. J Bacteriol 2003; 185:1:274-284.
- Bassler B., Wright M., Silverman M. Sistemi di segnalazione multipli che controllano l'espressione della luminescenza in Vibrio Harveyi: sequenza e funzione dei geni che codificano per una seconda via sensoriale. Mol Microbiol 1994; 13:273-286.
- Ji G., Beavis R., Novick R. Interferenza batterica causata da varianti peptidiche autoinducenti. Scienza 1997; 276:2027-2030.
- Lee S., Park S., Lee J., et al. I geni che codificano per l’enzima N-acyi omoserina che degrada il lattone sono diffusi in molte sottospecie di Bacillus thuringiensis. AppleEnviron Microbiol 2002; 68:8:3919-3924.
- Leadbetter J., Greenberg E. Metabolismo dei segnali di rilevamento del quorum dell'acilomoserina lattone mediante Paradosso di Variovorax. J Bacteriol 2000; 182:6921-6926.
- Hoang T., Schweizer H. Caratterizzazione di Pseudomonas aeruginosa enoil-acil-proteina trasportatrice reduttasi (Fabl): un bersaglio per l'antimicrobico triclosan e il suo ruolo nella sintesi del lattone omoserina acilato. Ibid 1999; 181:. 5489-5497.
- Pearson J., Delden C., Iglewski B. L'efflusso attivo e la diffusione sono coinvolti nel trasporto di Pseudomonas aeruginosa segnali da cellula a cellula. J. batteriolo. 1999; 181: 1203-1210.
- Manefield M., Welch M., Givskov G. et al. Le furanosi alogenate dell'alga rossa, Delisea pulchra, inibiscono la sintesi degli antibiotici carbapenemici e la produzione del fattore di virulenza esoenzimatico nel fitopatogeno Erwinia carotovora. FEMS Microbiol Lett 2001; 205:131-138.
- Dong Y., Wang L., Xu J. et al. Estinzione dell'infezione batterica dipendente dal quorum sensing da parte di una N-acil omoserina lattonasi. Natura. 2001; 411:813-817.
- Romanova Yu.M., Gintsburg A.L. Le citochine sono possibili attivatori della crescita di batteri patogeni. Vesti RAMN 2000; 1:13-17.
- Barcina I., Lebaron P., Vives-Rego J. Sopravvivenza di batteri alloctoni nei sistemi acquatici: un approccio biologico. FEMS Microbiol Ecol 1997; 23:1-9.
- Heim S., Lleo M., Bonato B. et al. Lo stato vitale ma non coltivabile e la fame sono diverse risposte allo stress Enterococcus faecalis, come determinato dall'analisi del proteoma. J Bacteriol 2002; 184:23: 6739-6745.
- Xu H., Roberts N., Singleton F. el al. Sopravvivenza e vitalità del non-culturale Escherichia coli E Vibrio cholerae nell'ambiente estuario e marino. Microb Ecol 1982; 8:313-323.
- Kell D., Kaprelyants A., Grafen A. Feromoni, comportamento sociale e funzioni del metabolismo secondario nei batteri. Tendenze nell'ecologia e nell'evoluzione 1995; 10:126-129.
- Domingue G., Woody H. Persistenza batterica ed espressione della malattia. Clin Microbiol Rev 1997; 10:320-328.
- Khomenko A. La variabilità di Mycobacterium tuberculosis in pazienti affetti da tubercolosi polmonare cavitaria in corso di chemioterapia. Malattia polmonare tubercolare 1987; 68:243-253.
- Gangadharam P. Dormienza micobatterica. Tub Lung Dis 1995; 76:477-479.
- Wayne L. Dormienza di Mycobacterium tuberculosis e latenza della malattia. Europeo J Clin Microbiol Infect Dis 1994; 13:908-914.
- Wayne L., Hayes L. An in vitro modello per lo studio sequenziale dello spostamento verso il basso di Mycobacterium tuberculosis attraverso 2 stadi di persistenza non replicante. Infetta Immuni 1996; 64:2062-2069.
- Beumer R., Devries J., Rombouts F. Campylobacter jejuni cellule coccoidi non coltivabili. Stagista J Food Microbiol 1992; 15:153-163.
- Kusters J., Gerrits M., Van Strijp J. el at. forme coccoidi di Helicobacter pylori sono le manifestazioni morfologiche della morte cellulare. Infetta Immun 1997; 65:3672-3679.
- Cellini L., Hui P., Leung K. el at. Coccoide Helicobacter pylori non coltivabile in vitro ritorna nei topi. Microbiol Immun 1994; 38:843-850.
- Hirsch C., Yoneda T., Averill L. et al. Miglioramento della crescita intracellulare di Mycobacterium tuberculosis nei monociti umani trasformando il fattore di crescita-b-l. J. Infect Dis 1994; 170:1229-1237.
- Bermudez. L., Pelrofsky M. Regolazione dell'espressione di Mycobacterium avium le proteine complesse differiscono a seconda dell'ambiente all'interno delle cellule ospiti. Immunol Cell Biol 1997; 75:35-40.
- Woods D., Jones A., Hill P. Interazione dell'insulina con Pseudomonas pseudomallei. Infetta Immun 1993; 61:4045-4050.
- Fortier A., Leiby D., Narayanan R. et al. Crescita di Francisella tularensis LVS nei macrofagi: il compartimento intracellulare acido fornisce il ferro essenziale necessario per la crescita. Ibid 1995; 65:1478-1483.
- Duncan S., Glover L., Killham K., Prosser J. Rilevamento basato sulla luminescenza dell'attività di batteri affamati e vitali ma non coltivabili. Apple Ambiente Microbiol 1994; 60:1308-1316.
- Young D., Duncan K. Prospettive per nuovi interventi nel trattamento e nella prevenzione delle malattie micobatteriche. Anna. Rev. microbiolo. 1995; 49:641-673.
- Mukamolova G., Kapreilyants A., Young D. et al. Una citochina batterica. Proc Nat! Acad.Sci USA. 1998; 95: 8916-8921.
- Shleeva M.O., Mukamolova G.V., Telkov M.V. et al.Formazione di cellule “incolte”. Mycobacterium tuberculosis e la loro rinascita. Microbiologia 2003; 72:76-83.
) comunicano e coordinano il loro comportamento attraverso la secrezione di segnali molecolari.
Assegnazione del senso del quorum
Lo scopo del quorum sense è coordinare determinati comportamenti o azioni tra batteri della stessa specie o sottospecie, a seconda della loro densità di popolazione. Ad esempio, batteri patogeni opportunisti Pseudomonas aeruginosa possono moltiplicarsi all'interno dell'ospite senza danni, purché non raggiungano una certa concentrazione. Ma diventano aggressivi quando il loro numero diventa sufficiente a sopraffare il sistema immunitario dell'ospite, portando allo sviluppo della malattia. Per fare ciò, i batteri devono formare biofilm sulla superficie del corpo dell'ospite. È possibile che la degradazione enzimatica terapeutica delle molecole di segnalazione prevenga la formazione di tali biofilm. La distruzione del processo di segnalazione in questo modo equivale alla soppressione del senso del quorum.
Il ruolo del quorum sensing in alcuni organismi
Il rilevamento del quorum è stato osservato per la prima volta nei batteri Vibrio fischeri, un batterio bioluminescente che vive come simbionte negli organi luminosi di una specie di calamaro hawaiano. Quando le cellule Vibrio fischeri vivono liberamente, gli autoinduttori sono in bassa concentrazione e quindi le cellule non sono luminescenti. Nell'organo luminoso del calamaro (fotofori) essi sono estremamente concentrati (circa 10 11 cellule/ml), e quindi viene indotta la trascrizione della luciferasi, portando alla bioluminescenza.
I processi che sono regolati o parzialmente regolati dal rilevamento del quorum basato su AI-2 in E. coli includono la divisione cellulare. In altre specie, ad esempio - Pseudomonas aeruginosa(Pseudomonas aeruginosa), i processi associati al rilevamento del quorum includono lo sviluppo di biofilm, la produzione di esopolisaccaridi e l'aggregazione cellulare. È stato scoperto che l'AI-2 aumenta l'espressione del gene sdiA, un regolatore trascrizionale del promotore che regola il gene ftsQ, parte dell'operone ftsQAZ importante per la divisione cellulare.
Streptococco pneumoniae(pneumococco) utilizza il rilevamento del quorum per rendere competenti le cellule. Ciò può essere importante per aumentare il numero di mutazioni in condizioni di sovrappopolazione, quando diventa necessario colonizzare nuovi ambienti.
Scrivi una recensione su "Quorum Sense"
Passaggio del quorum
Quella stessa notte, inchinandosi al ministro della Guerra, Bolkonskij si recò all'esercito, non sapendo dove l'avrebbe trovata e temendo di essere intercettato dai francesi sulla strada per Krems.A Brunn tutta la popolazione della corte fece le valigie e carichi pesanti furono già inviati a Olmutz. Vicino a Etzelsdorf, il principe Andrei si imboccò la strada lungo la quale l'esercito russo si muoveva con la massima fretta e nel più grande disordine. La strada era così affollata di carri che era impossibile viaggiare in carrozza. Prendendo un cavallo e un cosacco dal capo cosacco, il principe Andrey, affamato e stanco, superando i carri, andò a cercare il comandante in capo e il suo carro. Lungo la strada gli giunsero le voci più inquietanti sullo stato dell'esercito, e la vista dell'esercito che correva in disordine confermò queste voci.
"Cette armee russe que l" or de l "Angleterre a transportee, des extremites de l" univers, nous allons lui faire eprouver le meme sort (le sort de l "armee d" Ulm)", ["Questo esercito russo, che L'oro inglese è stato portato qui dalla fine del mondo, subirà lo stesso destino (il destino dell'esercito di Ulm). "] Ha ricordato le parole dell'ordine di Bonaparte al suo esercito prima dell'inizio della campagna, e queste parole ugualmente suscitò in lui sorpresa per l'eroe geniale, un sentimento di orgoglio offeso e speranza di gloria: "E se non restasse altro che morire? pensò. Ebbene, se necessario! Non lo farò peggio degli altri".
Il principe Andrei guardò con disprezzo queste infinite squadre che interferivano, carri, parchi, artiglieria e ancora carri, carri e carri di tutti i tipi possibili, che si sorpassavano l'un l'altro e bloccavano la strada fangosa in tre, quattro file. Da ogni parte, dietro e davanti, a perdita d'orecchio, rumore di ruote, rombo di corpi, carri e affusti di armi, scalpiccio di cavalli, colpi di frusta, grida di pungolii, imprecazioni di soldati, furono ascoltati attendenti e ufficiali. Lungo i bordi della strada si vedevano incessantemente cavalli caduti, scuoiati e non scuoiati, poi carri rotti, su cui sedevano, in attesa di qualcosa, soldati solitari, poi soldati separati dalle squadre, che in folla si dirigevano verso i villaggi vicini o trascinando dai villaggi polli, montoni, fieno o fieno, sacchi pieni di qualcosa.
Nelle discese e nelle salite la folla si faceva più fitta e si levava un gemito di grida ininterrotto. I soldati, annegando nel fango fino alle ginocchia, raccolsero armi e carri tra le braccia; le fruste battevano, gli zoccoli scivolavano, le tirelle scoppiavano e i petti scoppiavano di urla. Gli ufficiali incaricati del movimento, sia in avanti che all'indietro, passavano tra i convogli. Le loro voci si udivano appena in mezzo al rimbombo generale, e dai loro volti si vedeva che disperavano della possibilità di porre fine a quel disordine. “Voila le cher [‘Ecco un costoso] esercito ortodosso”, pensò Bolkonskij, ricordando le parole di Bilibin.
I titolari del brevetto RU 2534617:
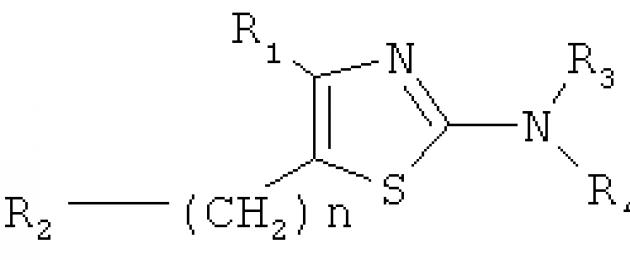
L'invenzione riguarda il campo della microbiologia, della biotecnologia e della farmaceutica, vale a dire piccole molecole regolatrici in grado di modificare la direzione della comunicazione dipendente dalla densità e del comportamento collettivo che regola ("quorum sense") nei batteri. In particolare, l'invenzione riguarda l'uso di un derivato tiazolico di formula 1 come regolatore (attivatore o inibitore) del comportamento collettivo ("quorum sensing") nei batteri. EFFETTO: derivato tiazolico destinato alla regolazione del "quorum sense" mediato dai lattoni omoserina nei batteri biotecnologicamente utili, putrefattivi e patogeni produttori di violaceina. 2 illustrato, 1 tab., 2 pr.
L'invenzione riguarda la microbiologia, la biotecnologia e la farmaceutica e riguarda piccole molecole regolatrici in grado di cambiare direzione (indebolire o rafforzare) la comunicazione dipendente dalla densità e il comportamento collettivo che regola ("quorum sense") nei batteri. L'invenzione può essere utilizzata nel controllo dei processi biotecnologici, nella produzione di mezzi per prevenire il deterioramento dei prodotti agricoli, nonché nella creazione di nuovi farmaci progettati per controllare e gestire le infezioni batteriche nelle piante, negli animali e nell'uomo.
La scoperta della comunicazione dipendente dalla densità nei batteri con la caratterizzazione dei meccanismi genetici molecolari sottostanti è stata una delle scoperte più sorprendenti della microbiologia della fine del XX secolo. Allo stesso tempo, questo fenomeno del comportamento collettivo dei batteri, designato dal concetto di "quorum sensing" (inglese - quorum sensing), ha consentito una valutazione fondamentalmente nuova di una serie di esempi di differenziazione funzionale e morfologica dei procarioti, incluso il sviluppo della bioluminescenza, sintesi di pigmenti e antibiotici, formazione di esoenzimi e fattori di virulenza, formazione di biofilm, coniugazione e sporulazione.
La prima delle varianti descritte e la più comune del "quorum sensing" tra i microrganismi sono i sistemi simili a luxI/luxR, in cui la molecola di segnalazione autoinduttrice sintetizzata sotto il controllo del gene luxI si diffonde nell'ambiente esterno e, una volta raggiunto il punto critico, densità di popolazione e la propria concentrazione soglia da questa determinata, inverte il movimento all'interno della cellula batterica, dove, legandosi alla proteina regolatrice LuxR, innesca la trascrizione dei geni bersaglio. Allo stesso tempo, l'analisi della natura chimica di tali autoinduttori ha permesso di caratterizzarli come varie varianti di omoserina lattoni acilati (HSL).
Decifrare i meccanismi genetici molecolari del comportamento collettivo, nonché identificare l’importante ruolo biologico dei sistemi di comunicazione dipendenti dalla densità, ha determinato l’importanza della ricerca di approcci per gestire il senso del quorum. Le soluzioni proposte erano: 1) soppressione della sintesi autoinduttrice; 2) la sua degradazione da parte di enzimi specifici (lattonasi o acilasi); 3) l'uso di agonisti e antagonisti del GSL capaci di interferire direttamente con il segnale naturale di legame alle proteine luxR-like. È proprio quest'ultimo approccio, che è quello sviluppato più intensamente in molti laboratori in tutto il mondo e che ha portato fino ad oggi alla creazione di diverse centinaia di composti attivi, che costituisce la base teorica per la presente invenzione.
Un'analisi delle fonti brevettuali aperte ci consente di affermare che l'analogo più vicino dell'invenzione rivendicata è un brevetto, la cui formula e descrizione contengono informazioni su un numero di composti, a seconda dell'oggetto dell'influenza, in grado di provocare l'attivazione (agonista ) o effetti inibitori (antagonisti) in relazione al "quorum sensing" mediato dai lattoni omoserina in alcuni tipi di batteri. Allo stesso tempo, i richiedenti hanno basato tali sostanze su un anello lattonico simile all'HSL naturale, per conferirgli ulteriori attività modulanti, è stata effettuata una modifica covalente con gruppi acilici di varie strutture e composizioni. Tuttavia, la significativa somiglianza strutturale del gruppo di molecole proposto con segnali naturali non solo fornisce la possibilità di interferenza tra loro indicata dai richiedenti, ma mantiene potenzialmente la possibilità di sviluppare effetti non spiegati in relazione ad altri microrganismi, comunicazione dipendente dalla densità tra che è mediato da GSL strutturalmente simili.
A sua volta, per quanto riguarda la struttura chimica del composto rivendicato, la soluzione tecnica conosciuta più vicina è un brevetto, la cui formula e descrizione contengono informazioni su un numero di composti basati su un anello tiazolico a cinque membri legato covalentemente a cicloalchile sostituito o non sostituito, arile e altri gruppi. Tuttavia, questo brevetto non indica la possibilità di utilizzare questi composti per regolare il comportamento collettivo (“quorum sensing”) nei batteri, e lo scopo principale dei composti rivendicati è il loro utilizzo come antagonisti dei recettori dell'adenosina.
Pertanto, l'invenzione rivendicata non è nota dallo stato della tecnica, il che ne determina la rispondenza al requisito di novità.
L'obiettivo della presente invenzione è sviluppare un composto strutturalmente diverso dalle lattine omoserina, che abbia una capacità selettiva e pronunciata di regolare (sia aumentare che diminuire) il comportamento collettivo mediato da GSL ("quorum sensing") in un certo intervallo di funzioni biotecnologicamente utili, batteri putrefattivi e patogeni.
Nella presente invenzione, questo problema viene risolto utilizzando un composto a base di tiazolo, completamente descritto dalla formula 1:
La presente invenzione descrive la formula strutturale del composto di formula 1 e i metodi della sua applicazione pratica per la regolazione del comportamento collettivo ("quorum sensing") nei batteri.
Secondo la presente invenzione, la preparazione (composizione) regolatoria a base di un derivato tiazolico contiene in peso dallo 0,0001 al 100% dei composti di formula 1, il resto sono componenti neutri o sostanze che modificano positivamente (aumentano la biodisponibilità, aumentano la durata di azione, ecc.) le proprietà di queste composizioni.
Rispetto ai composti che costituiscono l'essenza dei brevetti noti, il composto rivendicato di formula 1 presenta una serie di differenze significative, vale a dire:
In primo luogo, contrariamente ai composti noti basati sull'anello lattonico e, a questo proposito, essendo stretti analoghi strutturali delle molecole naturali autoregolatrici - lattoni omoserina, il composto rivendicato è un ligando sintetico strutturalmente diverso da essi. I regolatori "quorum sensing" a base di tiazolo non sono noti dalla letteratura scientifica e brevettuale disponibile;
in secondo luogo, contrariamente ai noti derivati tiazolici della formula generale 2

il composto rivendicato ha solo una variante del radicale aggiunto covalentemente, corrispondente a R 4 nella sua posizione e completamente descritta dal residuo di acido esanoico analogo alla molecola segnale naturale (esanoil-omoserina lattone), in assenza di altri cambiamenti nei radicali R 1 , R 2 e R 3 \u003d H . Inoltre, contrariamente al brevetto noto, che prevede l'uso di composti di formula generale 2 come antagonisti dei recettori dell'adenosina, il composto di formula 1 rivendicato è destinato a regolare il comportamento collettivo mediato da GSL ("quorum sensing") nei batteri;
in terzo luogo, a causa delle differenze strutturali rispetto alle molecole autoregolatrici naturali che sono attive in molti sistemi simili a luxI/luxR, il composto rivendicato di formula 1 ha un'attività regolatoria selettiva (selettiva) implementata in relazione al sistema di biosintesi della violaceina regolato da cviI/cviR Chromobacterium violaceum, e anche altri batteri produttori di violaceina utili dal punto di vista biotecnologico, putrefattivi e patogeni (vedi esempio 1). In questo caso, la probabile ragione dell'azione selettiva del composto di formula 1 nei cosiddetti sistemi "quorum sensing" risiede nell'interazione selettiva con la proteina regolatrice CviR e suoi omologhi stretti, ma non con altre proteine LuxR-like. A sua volta, la diversità naturale delle proteine simili a CviR determina la possibilità della loro regolazione sia positiva che negativa da parte del composto rivendicato, che in una diversa gamma di batteri produttori di violaceina biotecnologicamente utili, putrefattivi e patogeni si manifesterà sia come aumento ( vedi esempio 1) o come indebolimento (vedi esempio 2) del comportamento collettivo.
Per comprendere l'essenza dell'invenzione, è anche necessario sottolineare che la regolazione del "quorum sensing" ottenuta utilizzando il composto di formula 1 include, ma non è limitata a, l'effetto sulla produzione di violaceina, poiché Sotto il controllo della proteina regolatrice CviR e dei suoi omologhi, ci sono una serie di geni bersaglio (operoni), compresi quelli responsabili della produzione di esoenzimi e della formazione di biofilm. L'uso del test per l'induzione o l'inibizione della biosintesi della violaceina nella presente invenzione è determinato dalla semplicità e dal contenuto informativo della manifestazione registrata dell'attività regolatoria del composto di formula 1.
Pertanto, il risultato dell'azione del composto di formula 1 è la regolazione specifica di un certo sistema di "quorum sensing", la cui direzione (intensificazione o indebolimento) è determinata dalle caratteristiche recettoriali delle proteine CviR-simili che percepiscono Esso. Pertanto, utilizzando lo stesso composto, è possibile influenzare il comportamento collettivo di vari microrganismi in direzioni diverse, anche isolatamente o nella loro coltura mista.
L'uso protetto del composto di formula 1 implica, tra le altre cose, il suo utilizzo per il controllo di processi biotecnologici attuati con l'aiuto di microrganismi produttori di violaceina (per riferimento: la violaceina è un derivato indolico formato durante l'ossidazione del triptofano, un composto blu -pigmento viola con attività antibatterica, protistocida, antivirale ed altre attività biotecnologicamente e farmacologicamente utili). In questo caso, il composto di formula 1 può essere introdotto in mezzi nutritivi solidi o liquidi sotto forma di soluzioni, nonché utilizzato come sostanza pura o immobilizzato su vari supporti.
L'invenzione brevettata prevede anche l'uso di un composto di formula 1 per regolare l'attività di altri geni bersaglio (operoni), compresi quelli coinvolti nel deterioramento dei prodotti agricoli, nonché nello sviluppo di malattie infettive di piante, animali e esseri umani. A tal fine, questo composto può essere somministrato all'organismo per fornire un effetto sistemico, nonché applicato localmente per influenzare determinate aree (ad esempio come parte di medicazioni per ferite, quando si tratta il campo chirurgico, ecc.). Il composto può essere utilizzato sotto forma di solidi, soluzioni o sospensioni in acqua o altri solventi e applicato a una varietà di veicoli. È anche possibile utilizzare un composto di formula 1 in composizioni con altre sostanze, inclusa la modifica positiva (aumento della biodisponibilità, durata dell'azione) della sua attività biologica.
L'invenzione rivendicata è illustrata, ma non limitata in alcun modo, dai seguenti esempi.
Esempio 1. Stimolazione del comportamento collettivo ("quorum sensing") nei batteri.
La determinazione della capacità dei composti di formula 1 di regolare il "quorum sense" è stata effettuata utilizzando due sistemi di test batterici, in presenza di esanoil-omoserina lattone (C 6 -GSL)? responsabile della sintesi del pigmento violaceina (Chromobacterium violaceum NCTC 13274) o dello sviluppo della bioluminescenza (Escherichia coli pAL103). Allo stesso tempo, la prima caratteristica è stata l'inserimento del trasposone Tn5 nel gene cvil, che è responsabile della sintesi del proprio C 6 -HSL, pur mantenendo il gene cviR funzionalmente attivo e la proteina regolatrice da esso codificata, che è responsabile della percezione dell'autoinduttore.
Quomm sensing e Chrornobacteriurn violaceum: sfruttamento della produzione di violaceina e inibizione per il rilevamento dei lattoni N-acil omoserina. Microbiologia, 1997, V.143, P.3703-3711]. A sua volta, una caratteristica del secondo ceppo era la presenza del costrutto genetico luxR + luxI_luxCDABE, che codifica per la proteina recettore LuxR di Vibrio fischeri e, in presenza di C 6 -GSL o C 6 -oxo-GSL introdotti esogenamente, responsabile della sviluppo della luminescenza (bioluminescenza).
Durante i test, C. violaceum NCTC13274 ed Escherichia coli pAL103 sono stati coltivati su terreni nutritivi liquidi in assenza (controllo) e in presenza di C 6 -GSL o di un composto di formula 1 (esperimento) utilizzato nell'intervallo di concentrazione da 2 a 1000 µm. La caratteristica dell'azione regolatoria era il valore EC50 - la concentrazione dei composti confrontati, che provoca l'induzione della formazione del pigmento violaceina o della bioluminescenza del 50% dell'effetto più pronunciato in presenza di un segnale naturale. I risultati di tali test sono illustrati nella Figura 1 e sono riepilogati nella Tabella 1.
Tabella 1. Valutazione dell'effetto del composto di formula 1 sul comportamento collettivo ("quorum sensing") nei batteri nei test per C. violaceum NCTC13274 ed E. coli pAL103.
Dai dati presentati si può vedere che entrambi i microrganismi utilizzati reagiscono intensamente con la sintesi quorum-dipendente della violaceina (C. violaceum NCTC) o con lo sviluppo della bioluminescenza (E. coli pAL103) in presenza dell'autoregolatore naturale C 6 -GSL . A sua volta, il composto in esame di formula 1 agisce in modo meno attivo, ma più specifico, provocando l'induzione della sintesi della violaceina, ma non lo sviluppo della bioluminescenza. Allo stesso tempo, queste differenze sono presumibilmente basate sull'affinità selettiva del composto 1 per la proteina CviR che riceve il segnale regolatorio, in assenza di tale affinità per LuxR.
Un risultato positivo di tale utilizzo dell'invenzione rivendicata è la possibilità di induzione selettiva del "quorum sensing" di alcuni tipi di batteri che fanno parte di associazioni polimicrobiche.
Esempio 2. Soppressione del comportamento collettivo (“quorum sensing”) nei batteri.
La determinazione della capacità dei composti di formula 1 di regolare il "senso del quorum" è stata effettuata utilizzando un ceppo di Jantinobacterium lividum depositato nella Collezione tutta russa di microrganismi industriali (VKPM) con il numero B-10136. Questo ceppo è un isolato naturale caratterizzato dalla capacità di sintetizzare il pigmento violaceina sotto il controllo di un autoinduttore non identificato.
Durante il test, J. lividum B-10136 è stato coltivato su terreni nutritivi liquidi in assenza (controllo) e in presenza del composto di formula 1 (esperimento), utilizzato nell'intervallo di concentrazione da 2 a 1000 μm. La caratteristica dell'azione regolatoria era il valore di EC50 - la concentrazione del composto di formula 1, che causava la soppressione della produzione di violaceina del 50% dell'effetto più pronunciato nel controllo.
I risultati di tali test sono illustrati in Fig.2. Dai dati sopra riportati, ne consegue che il composto di prova di formula 1 inibisce la produzione di violaceina (EC50=87,5 μM), che lo caratterizza come un inibitore del comportamento collettivo (“quorum sensing”) di J. lividum B-10136.
Un risultato positivo di tale utilizzo dell'invenzione rivendicata è la capacità di sopprimere il "quorum sense" di alcuni tipi di batteri, in particolare J. lividum, per prevenire i danni da essi causati ai prodotti agricoli. La stessa attività può essere utilizzata nel trattamento e nella prevenzione delle malattie infettive delle piante, degli animali e dell'uomo causate da J. lividum e altri microrganismi produttori di violaceina.
L'uso di un derivato tiazolico di formula 1 come regolatore (attivatore o inibitore) del comportamento collettivo ("quorum sensing") nei batteri: 
Brevetti simili:
L'invenzione riguarda il campo della chimica organica, vale a dire nuove sostanze biologicamente attive della classe dei 4-aril-2-idrossi-4-osso-2-butenoati di eterolammonio, vale a dire il 2-idrossi-4-metilfenil-4- formula dell'osso-2-butenoato tiazolinil ammonio (1).
L'invenzione riguarda il campo della chimica organica, nuove sostanze biologicamente attive della classe dei 4-aril-2-idrossi-4-osso-2-butenoati di eterilammonio, vale a dire 2-idrossi-4-osso-4-(4 -clorofenil)-2 -tiazolina ammonio butenoato (1) della formula avente attività anticoagulante, che ne suggerisce l'uso in medicina come agente anticoagulante.
L'invenzione riguarda nuovi derivati delle 2-(imminosostituite) tiazolidine, un metodo per la loro preparazione, prodotti farmaceutici contenenti queste sostanze, l'uso di questi derivati delle 2-(imminosostituite) tiazolidine per il trattamento di varie malattie, nonché come la produzione di composizioni farmaceutiche basate su di essi, utilizzate per il trattamento.
L'invenzione riguarda un metodo per l'ottenimento di nuovi composti chimici biologicamente attivi, in particolare un metodo per l'ottenimento di nuovi derivati imminotiazolidinici o loro cloridrati ad attività antidepressiva, antiepilettica, antiparkinsoniana ed analgesica.
L'invenzione riguarda un composto rappresentato dalla formula (I), dove rappresenta la configurazione α; è una configurazione β; e rappresenta la configurazione α, la configurazione β, o qualsiasi loro miscela, il suo sale, o la sua miscela con un diastereomero in un rapporto arbitrario, o il suo clatrato di ciclodestrina.
L'invenzione riguarda un metodo per ottenere cristalli della forma A dell'acido 2-(3-ciano-4-isobutilossifenil)-4-metil-5-tiazolocarbossilico. Il metodo comprende: la fase di dissoluzione dell'acido 2-(3-ciano-4-isobutilossifenil)-4-metil-5-tiazolocarbossilico in 1-propanolo o 2-propanolo mediante riscaldamento, la fase di raffreddamento della soluzione risultante e la fase di aggiungere eptano a questa soluzione.
L'invenzione riguarda l'uso di composti di formula generale (I) aventi le proprietà di inibitore delle monoaminossidasi (MAO) e/o di perossidazione lipidica, e/o le proprietà di modulatori dei canali del sodio, nonché un farmaco a base di essi , avendo le stesse proprietà, più specificatamente i composti e il farmaco possono essere usati per trattare il morbo di Parkinson, la demenza senile, il morbo di Alzheimer, la corea di Huntington, la sclerosi laterale amiotrofica, la schizofrenia, la depressione, la psicosi, il dolore e l'epilessia.
L'invenzione riguarda un composto rappresentato dalla formula (I) in cui Al è benzene o un eterociclo scelto dal gruppo costituito da piridina, pirazina, imidazolo, tiazolo, pirimidina, tiofene, piridazina, benzossazina e ossobenzossazina; A2 è benzene eventualmente sostituito con fluoro o tiofene; B1 è idrogeno, alchile inferiore facoltativamente sostituito con piperazinile o morfolino, alchile inferiore alogenato, alcossi inferiore sostituito con carbamoile, acilammino, carbamoile o alchilcarbonilossi inferiore (a condizione che quando A1 è tiazolo, B1 non è acilammino); B2 è idrogeno o un gruppo funzionale contenente almeno un atomo di azoto selezionato dal gruppo costituito da acilammino, pirrolidinile, morfolino, piperidinile facoltativamente sostituito con acile, piperazinile facoltativamente sostituito con alchile inferiore o acile, pirazolile, diazabicicloeptile, facoltativamente sostituito con acile, e di-(alchil)ammino inferiore, ammino o acilammino facoltativamente sostituito (a condizione che quando A1 sia tiazolo, B2 non sia acilammino); Y è un gruppo rappresentato dalla formula (II) in cui J è etilene o alchinilene inferiore; L è un legame; M sta per legame; X è -(CH2)m-, -(CH2)m-O- o -(CH2)m-NR2- (dove m è un numero intero da 0 a 3 e R2 è idrogeno); D è -NR3- dove R3 è idrogeno; ed E è un ammino, o un suo sale farmaceuticamente accettabile.
L'invenzione riguarda composti di formula 1.0: dove Q è un anello tetraidropiridinilico sostituito. R5, R1 sono selezionati dal gruppo costituito da: (1) piridile sostituito con un sostituente selezionato dal gruppo costituito da: -O-CH3, -O-C2H5, -O-CH(CH3)2 e -O-( CH2 )2-O-CH3, R2 è selezionato dal gruppo costituito da: -OCH3 e -SCH3; e R5 è selezionato dal gruppo costituito da: (a) triazolilfenil- sostituito, dove triazolile è sostituito con uno o due gruppi alchilici selezionati dal gruppo costituito da: -C1-C4 alchile, (b) triazolilfenil- sostituito, dove triazolile è sostituito sull'atomo di azoto -C1-C4 alchile, (c) triazolilfenil- sostituito, dove triazolile è sostituito sull'atomo di azoto -C2 alchilene-O-C1-C2 alchile, (d) triazolilfenil- sostituito, dove triazolile è sostituito sull'atomo atomo di azoto -C2-C4 alchilene-O-CH3, e (e) triazolilfenil- sostituito, in cui il triazolile è sostituito sull'atomo di azoto con -C1-C4 alchile idrossi-sostituito, e in cui il fenile è facoltativamente sostituito con da 1 a 3 sostituenti selezionati indipendentemente dal gruppo costituito da alogeno; e i loro sali e solvati farmaceuticamente accettabili, che si ritiene siano inibitori di ERK.
L'invenzione riguarda un nuovo agente, che è un derivato della rodanina di formula (I), per il trattamento di malattie tumorali di varia localizzazione. EFFETTO: mezzo di azione antiproliferativa e antimetastatica per il trattamento delle malattie tumorali.
L'uso di (R)-5--2-(-propylimino)-3-orto-toliltiazolidin-4-one (Composto 1) o del suo sale per la preparazione di un medicinale per la prevenzione e/o il trattamento di una malattia o viene proposto un disturbo associato all'attivazione del sistema immunitario.sistemi in cui il farmaco è un insieme di dosi del Composto 1, e durante la fase iniziale del trattamento, la dose induce desensibilizzazione cardiaca ed è inferiore alla dose finale, e durante il periodo specificato nella fase iniziale del trattamento, la dose viene somministrata ad una frequenza che mantiene la desensibilizzazione cardiaca fino al successivo brusco calo della frequenza cardiaca, e quindi la dose viene titolata fino alla dose finale del Composto 1; metodo di trattamento e serie di dosi appropriati.
L'invenzione riguarda un composto di formula I o suoi sali terapeuticamente accettabili, dove A1 è furile, imidazolile, isotiazolile, isossazolile, pirazolile, pirrolile, tiazolile, tiadiazolile, tienile, triazolile, piperidinile, morfolinile, diidro-1,3,4- tiadiazol-2-il, benzotien-2-il, benzotiazol-2-il, tetraidrotien-3-il, triazolopirimidin-2-il o imidazo-tiazol-5-il; dove A1 non è sostituito o sostituito con uno o due o tre o quattro o cinque sostituenti selezionati indipendentemente da R1, OR1, C(O)OR1, NHR1, N(R1)2, C(N)C(O)R1, C( O)NHR1, NHC(O)R1, NR1C(O)R1, (O), NO2, F, Cl, Br e CF3; R1 è R2, R3, R4 o R5; R2 è fenile; R3 è pirazolile o isossazolile; R4 è piperidinile; R5 è alchile C1-C10 o alchenile C2-C10, ciascuno non sostituito o sostituito con sostituenti selezionati tra R7, SR7, N(R7)2, NHC(O)R7, F e CI; R7 è R8, R9, R10 o R11; R8 è fenile; R9 è ossadiazolile; R10 è morfolinile, pirrolidinile o tetraidropiranile; R11 è alchile C1-C10; Z1 è fenilene; Z2 è piperidina, non sostituita o sostituita con OCH3, o piperazina; Mancano entrambi Z1A e Z2A; L1 è alchile C1-C10 o alchenile C2-C10, non sostituito o sostituito con R37B; R37B è fenile; Z3 è R38 o R40; R38 è fenile; R40 è cicloesile o cicloesenile; dove il fenilene rappresentato da Z1 è non sostituito o sostituito da un gruppo OR41; R41 è R42 o R43; R42 è fenile che non è fuso o è fuso con pirrolile, imidazolile o pirazolo; R43 è piridinile che non è fuso o fuso con pirrolile; dove ciascuna delle porzioni cicliche di cui sopra rappresentate da R2, R3, R4, R8, R9, R10, R38, R40, R42 e R43 è indipendentemente non sostituita o sostituita con uno o più sostituenti selezionati indipendentemente da R57, OR57, C(O)OR57 , F, CI CF3 e Br; R57 è R58 o R61; R58 è fenile; R61 è alchile C1-C10; e dove il fenile rappresentato dal gruppo R58 non è sostituito o sostituito con uno o più sostituenti selezionati indipendentemente da F e CI. L'invenzione riguarda inoltre una composizione farmaceutica contenente tali composti, e un metodo per il trattamento di malattie in cui sono espresse le proteine anti-apoptotiche Bcl-2. 4 n. e 3 z.p. f-ly, 2 tavole, 48 pr.
L'invenzione riguarda il campo della microbiologia, della biotecnologia e della farmaceutica, vale a dire piccole molecole regolatrici in grado di modificare la direzione della comunicazione dipendente dalla densità e il comportamento collettivo da essa regolato nei batteri. In particolare l'invenzione riguarda l'uso di un derivato tiazolico di formula 1 come regolatore del comportamento collettivo nei batteri. EFFETTO: derivato tiazolico destinato alla regolazione del "quorum sense" mediato dai lattoni omoserina nei batteri biotecnologicamente utili, putrefattivi e patogeni produttori di violaceina. 2 illustrato, 1 tab., 2 pr.
La formazione, la crescita e la migrazione delle forme cellulari planctoniche per la colonizzazione nei biofilm sono regolate a livello di popolazione attraverso i meccanismi di comunicazione intercellulare. Il "quorum sensing" (QS) è un processo di coordinazione collettiva dell'espressione genica in una popolazione batterica che media il comportamento cellulare specifico. Il meccanismo di funzionamento del QS si basa su una complessa regolazione gerarchica dei loci bersaglio del genoma della cellula batterica. In questo caso, la regolazione viene effettuata a diversi livelli di influenza: trascrizionale, traduzionale, post-traduzionale.
Le cellule di una popolazione rispondono a uno specifico segnale cellulare con una risposta specifica. Ad oggi, è stato stabilito che le relazioni cellula-cellula influenzano la differenziazione cellulare intrapopolazione, l'espressione dei geni di virulenza, regolano i processi di crescita, la natura e la direzione della mobilità (tassi), nonché l'apoptosi batterica e la formazione di tossine.
Il lavoro di QS può essere paragonato alla regolazione ormonale dell'attività funzionale di vari organi e tessuti in un organismo multicellulare.
I microrganismi Gram-positivi e Gram-negativi utilizzano diversi sistemi di segnalazione e diversi trasmettitori di segnali chimici. I primi sintetizzano peptidi a 7-8 membri (Enterococcus spp.), ciclopeptidi (Staphylococcus spp.); secondo: una varietà di lattoni acil-omoserina (AHL).
Consideriamo il lavoro di QS sull'esempio di Pseudomonas aeruginosa. Questo microrganismo ha almeno tre sistemi regolatori. Il più studiato è il sistema LasI - LasR (AHL con una lunga catena acilica funge da segnale chimico); Sistema RhlI - RhlR (messaggero - AHL con una catena acilica corta, C4-HSL); e il sistema PQS dei chinoloni. L'interazione di questi tre sistemi permette di regolare l'espressione di circa il 6-10% del genoma. Nel sistema LasI - LasR, l'AHL sintasi, un prodotto del gene lasI, è responsabile della biosintesi delle molecole di segnalazione. La sua espressione è a livello basale, quindi l'accumulo di molecole segnale richiede molto tempo e l'effetto biologico inizia a manifestarsi solo nella fase stazionaria della crescita della popolazione. Nelle cellule, l'AHL interagisce con la proteina LasR (un prodotto del gene lasR, la cui espressione è anche a livello basale), formando un omodimero, un regolatore della trascrizione. Questo regolatore attiva molti geni coinvolti nella formazione della virulenza e nei processi di formazione del biofilm, attiva anche il regulon cromosomico las Box, che è responsabile dell'espressione di vari fattori di patogenicità (proteasi, elastasi, ecc.). Il complesso LasR + AHL attiva il secondo sistema di segnalazione. Ciò si verifica dopo l'interazione con il promotore del gene Rhl. L'espressione di RhlI provoca la formazione di una proteina per la sintesi di AHL con brevi residui acilici (C4-HSL). Il gene rhlR codifica per una proteina (RhlR) che interagisce con le molecole di segnalazione C4-HSL. Il risultante tandem proteico RhlR + C4-HSL regola la trascrizione dei geni che codificano per vari composti strutturali della matrice del biofilm (alginato, ramnolipide, ecc.), nonché lipasi e piocianina. Inoltre, questo regolatore trascrizionale attiva l'espressione di un altro regolatore - RpoS (fattore sigma della fase di crescita stazionaria di P. aeruginosa), che avvia la formazione di proteine dello stress nella cellula e partecipa a reazioni adattative. Tra gli isolati clinici di P.aeruginosa, è stato riscontrato che oltre al funzionamento dei sistemi di segnalazione AHL, entra in parallelo il sistema dei chinoloni (gene locus - pqsABCDE), i messaggeri sono idrossialchilchinoloni e idrossieptilchinoloni. Questo sistema funziona allo stesso modo dei meccanismi regolatori sopra descritti e media un aumento dell'espressione dei fattori di virulenza, in particolare la sintesi di elastasi e lectine. L'interazione dei tre sistemi di segnalazione colpisce un gran numero di geni, in relazione ai quali esiste una regolazione globale della trascrizione, che porta ad una labilità molto flessibile dei processi fisiologici della cellula, ed è una conseguenza dell'enorme potenziale adattativo di batteri nella popolazione.
I sistemi di segnalazione funzionano secondo il principio dell'autoinduzione, le molecole di segnalazione sintetizzate agiscono sulla propria cellula e, man mano che si accumulano nell'ambiente extracellulare, vengono attivati promotori sempre più dipendenti, i reguloni del genoma cellulare. Il QS basato su AHL è stato trovato in molti batteri Gram-negativi: Acinetobacter, Aeromonas, Brucella, Burkholderia, Erwinia, Enterobacter, Chromobacterium, Hafnia, Serratia, Vibrio, Yersinia, ecc. Strutture della molecola di segnalazione stessa.
Ma tra gli isolati clinici di batteri gram-negativi si osserva spesso anche la comunicazione incrociata, che garantisce l'interazione di popolazioni di diverse specie nel focus infettivo. Cross-QS può sia attivare che inibire i geni bersaglio dipendenti nelle associazioni batteriche. Ad esempio, P. aeruginosa, Serratia liquefaciens, Aeromonas hydrophila sintetizzano un tipo di molecole di segnalazione. Il QS di C.violaceum e A.idrophila è inibito dalle molecole AHL con lunghi residui acilici, che sono sintetizzati da vari microrganismi gram-negativi. Pseudomonas aeruginosa forma molecole di segnalazione con residui acilici lunghi e corti e non sono inibite a vicenda, tuttavia, i messaggeri di E. coli della stessa struttura molecolare con residui acilici lunghi sono in grado di inibire il sistema di segnalazione rhl di P. aeruginosa. Nei biofilm misti di P.aeruginosa e Burkholderia cepacia, Burkholderia risponde ai segnali di Pseudomonas aeruginosa (che a sua volta è insensibile ai segnali di B.cepacia), pertanto la popolazione di P.aeruginosa regola molti processi fisiologici del suo associato. Esistono prove che alcuni ceppi di P. aeruginosa isolati da pazienti con iscidosi non sono in grado di sintetizzare autonomamente gli autoinduttori del sistema di segnalazione rhl, con conseguente diminuzione della virulenza e formazione incompleta di biofilm negli esperimenti in vitro. Tuttavia, in vivo, questi stessi ceppi di Pseudomonas aeruginosa formano biofilm completi. Si è scoperto che la microflora isolata dal muco degli stessi pazienti sintetizza autoinduttori rhl, regolando così la virulenza e la formazione di biofilm di P.aeruginosa e avviando il processo infettivo. Le stesse molecole AHL influenzano diversamente altri gruppi di batteri; Le molecole di segnalazione dei procarioti possono anche influenzare il comportamento delle cellule fungine, vegetali e persino animali. Pertanto, P.aeruginosa AHL sopprime il processo di filamentazione della Candida albicans.
Negli esseri umani, le molecole AHL inibiscono la proliferazione dei leucociti e la formazione del fattore di necrosi tumorale b. Ad alte concentrazioni, l'AHL avvia l'apoptosi di vari tipi di cellule immunocompetenti. In generale, gli autoinduttori batterici hanno un effetto immunosoppressivo. È a causa delle reazioni QS che si instaurano relazioni “sociali” all'interno della popolazione, si forma una “rete di comunicazione chimica” di un biofilm, che può coprire una comunità multi-acqua.
Non meno interessante è il lavoro dei sistemi di segnalazione tra i microrganismi gram-positivi. Ad esempio, Enterococcus spp. QS regola il trasferimento dei plasmidi (dalle cellule del donatore alle cellule riceventi) attraverso il meccanismo della coniugazione. La cellula ricevente sintetizza un segnale peptidico specifico (feromone batterico “sessuale”), che si accumula nel mezzo e si lega specificamente ai recettori delle cellule donatrici che trasportano un plasmide che corrisponde a questo feromone. Il sistema di regolamentazione lanciato in questo caso garantisce l'espressione di fattori che mediano l'interazione cellulare e il trasferimento del plasmide (componenti di coniugazione). Come notato sopra, un feromone specifico corrisponde a un determinato plasmide. A causa di un meccanismo di interazione così rigido, viene effettuata la selezione batterica delle cellule all'interno del biofilm. Attraverso tale comunicazione, vengono traslocati plasmidi che trasportano geni per la resistenza agli antibiotici, geni per l'emolisina e batteriocine. Tipicamente, i peptidi segnale biologicamente attivi sono codificati nel cromosoma e le proteine recettrici che forniscono affinità per i feromoni sono codificate nei plasmidi stessi. Dopo la traslocazione del plasmide nella cellula ricevente, inizia la sintesi degli inibitori dei feromoni, ogni tipo di feromone ha il proprio inibitore. Questa funzione consente di disattivare il segnale per un plasmide esistente e migliorare l'accumulo di molecole di feromoni per un altro tipo di plasmide. cellula di microrganismi del biofilm
A causa del funzionamento di un tale sistema nella popolazione del biofilm, si verifica costantemente una selezione positiva di ceppi con proprietà benefiche e una selezione negativa - l'eliminazione di ceppi con fenotipi "non necessari". Nelle lesioni infettive, tali meccanismi di comunicazione per il trasferimento di elementi genetici mobili consentono di distribuire con la massima velocità i geni di resistenza agli antibiotici, virulenza e capacità fisiologiche aggiuntive.
Di grande interesse è il QS, che è coinvolto nella regolazione dell'espressione dei fattori di virulenza negli stafilococchi. La base genetica di questo sistema è agrBCD, il locus cromosomico. Come trasmettitori di segnali agiscono i ciclopeptidi: autoinduttori (AIP, peptide autoinducente), che sono classificati per struttura ed effetto biologico in gruppi e sottogruppi, ad esempio i sottogruppi 1 e 4 in S. aureus aumentano l'espressione dei fattori di virulenza. Queste molecole sono estremamente specifiche, la sostituzione anche di un solo amminoacido nella struttura del composto porta alla perdita della funzione biologica. Come negli esempi del sistema segnale-inibitore negli enterococchi, il sistema stafilococcico risponde solo a un tipo di autoinduttori, non appena la cellula ha ricevuto un segnale specifico, i geni inibitori vengono attivati e la cellula non è più in grado di percepire altri segnali . Questo meccanismo prevede una rigida selezione della popolazione. Le molecole segnale sintetizzate interagiscono con il sistema di membrana dell'istidina chinasi (agrC), che attiva il regolatore della trascrizione (agrA) attraverso una cascata di reazioni. Questa proteina regola bifunzionalmente i due promotori P2 e P3. Di conseguenza, le trascrizioni di questi geni dipendenti sono RNA II e RNA III, il primo contiene i principali geni agr, quindi si manifesta la risposta autoinduttiva del sistema. A sua volta, l'RNA III provvede alla regolazione della sintesi dei fattori di virulenza (DNasi, fibrinolisina, enterotossina, b-, c-, d-tossine, ecc.). Una caratteristica interessante in questa fase della regolazione è che la trascrizione dell'RNA III da 500 bp non contiene informazioni codificate, ad eccezione di un frame di lettura aperto per la d-tossina. La stragrande maggioranza della molecola di trascrizione stessa agisce come un inibitore ribosomiale. L'RNA III blocca il processo di traduzione del fattore di repressione della virulenza Rot (repressore delle tossine), che regola la sintesi delle tossine stafilococciche, con conseguente formazione incontrollata di esotossine. Pertanto, il sistema agr fornisce la regolazione della popolazione dell'espressione dei fattori di virulenza stafilococcica. Utilizzando varie varianti di studi PCR, si è scoperto che l'espressione dell'agr-locus nelle cellule è osservata in molte lesioni da stafilococco: infezioni cutanee, endocardite, artrite, sepsi. La popolazione del biofilm accumula molecole di segnalazione sintetizzate dalla stragrande maggioranza delle cellule, che costituiscono il "nucleo, quorum" metabolico e genetico della popolazione, determinano il comportamento metabolico e i cambiamenti fenotipici per tutte le cellule. Ciò è dovuto all'accumulo di segnali attraverso la proprietà di autoinduzione, e all'inibizione di altri segnali sintetizzati da una minoranza, o in generale da altri ceppi nel biofilm, grazie ad un meccanismo parallelo di inibizione. 1.5 Significato clinico dei biofilm.
Il concetto di biofilm, confermato utilizzando le moderne tecniche di imaging, ha cambiato il modo in cui guardiamo alle malattie infettive. Tutti i nuovi dati indicano che le infezioni croniche sono fondamentalmente diverse da quelle acute nella formazione di biofilm e che i fagociti del macroorganismo non sono in grado di assorbire i biofilm, a differenza delle singole cellule batteriche.
L’esistenza di biofilm nelle infezioni croniche richiede approcci completamente nuovi alla loro diagnosi e trattamento. Inoltre, i metodi batteriologici tradizionali non rilevano la maggior parte dei batteri coinvolti nel processo infettivo. Gli ultimi metodi molecolari, genomici, trascrizionali e proteomici hanno permesso di determinare che solo circa l'1% delle cellule di microbiocenosi patogene vengono determinate isolando una coltura pura. Di conseguenza, il trattamento prende di mira solo 1-2 specie batteriche tra i numerosi ceppi presenti nel biofilm (compresi possibilmente i funghi).
Ad oggi, il ruolo dei biofilm microbici nella comparsa e nello sviluppo di malattie comuni come le infezioni associate al cateterismo vascolare causate da Staphylococcus aureus e altri microrganismi Gram-positivi è stato dimostrato in modo affidabile; infezioni delle valvole cardiache e delle protesi articolari causate da stafilococchi; parodontite causata da un numero di microrganismi orali; infezioni delle vie urinarie determinate da E. coli e altri agenti patogeni; infezioni dell'orecchio medio - causano, ad esempio, Haemophilus influenzae, fibrosi cistica causata da P. aeruginosa, ecc.
Tutte queste malattie sono difficili da trattare, hanno un alto tasso di recidiva e alcune di esse possono essere fatali. I meccanismi attraverso i quali i microrganismi che formano biofilm causano processi patologici nel macroorganismo sono tutt’altro che chiari.
Oltre ai tessuti ospiti, i biofilm microbici colonizzano diversi dispositivi medici non biologici introdotti nel corpo umano (cateteri, pacemaker, valvole cardiache, dispositivi ortopedici). Studi su dispositivi medici impiantati mediante microscopia elettronica hanno dimostrato la presenza di biofilm batterici.
L’aumento della resistenza agli antibiotici e lo sviluppo di biofilm batterici rappresentano le principali sfide nel trattamento delle infezioni del tratto urinario.
È stato stabilito che le proprietà delle cellule e della matrice extracellulare sono alla base dell'aumento della resistenza. La matrice del biofilm può o meno legarsi e/o inattivare gli antibiotici. La resistenza dovuta alle proprietà delle cellule del biofilm si spiega con una diminuzione della loro superficie libera dovuta ai contatti tra loro e alla formazione di batteri speciali, chiamati persistenti.
Le persistenti sono cellule altruistiche che si formano nella fase stazionaria della crescita, sono metabolicamente inattive e garantiscono la sopravvivenza della popolazione materna in presenza di fattori letali per tutte le cellule. Nei biofilm, questa sottopopolazione costituisce l'1–5% della massa cellulare totale. La formazione di tali cellule dipende dal grado di crescita della popolazione; nella fase logaritmica la coltura non forma o forma una percentuale molto piccola di persistenti, il loro numero aumenta verso la fase stazionaria. La formazione di una sottopopolazione dipende inversamente dal livello di attività metabolica di tutte le cellule del biofilm, nonché dall'azione di fattori sfavorevoli esogeni. Il fenotipo persistente è caratterizzato da una biologia interessante; rallentano tutti i processi fisiologici e diventano tolleranti all'azione di vari fattori, compresi gli effetti dei farmaci antimicrobici.
La proprietà della tolleranza agli antibiotici differisce dai meccanismi di resistenza. L'azione di tutti i meccanismi di resistenza batterica, in sostanza, può essere ridotta a un fenomeno: questa è la prevenzione dell'interazione di un antibiotico con il suo bersaglio (a causa di cambiamenti nei bersagli stessi o attraverso la sintesi di enzimi che neutralizzano gli antibiotici ). La tolleranza è mediata dalla capacità di una cellula microbica di sopravvivere in presenza di un antibiotico rallentando il metabolismo e “spegnendo” i principali processi biologici della cellula.
I principali meccanismi di aumento della resistenza batterica agli antibiotici nei biofilm sono:
1. limitare la penetrazione degli antibiotici attraverso i biofilm;
2. la restrizione alimentare e il microambiente alterato nel biofilm portano ad una diminuzione del tasso di divisione batterica, per cui ci sono meno bersagli per l'azione degli antibiotici;
3. reazioni adattative;
4. variabilità genetica nei batteri che persistono in un biofilm.
Sulla base dei dati accumulati, ne consegue che gli antibiotici sono divisi in due tipi in base all'azione sui batteri nei biofilm. Il primo comprende gli antibiotici che penetrano nei biofilm e inibiscono o uccidono i microrganismi che li formano. Il secondo tipo sono gli antibiotici, che praticamente non penetrano nei biofilm, ma ne impediscono efficacemente l'insediamento a causa della migrazione dei batteri. Pertanto, alcuni antibiotici non penetrano nei biofilm e non distruggono le comunità esistenti, ma impediscono solo il loro aumento in numero e distribuzione nel corpo umano. A questo proposito, negli ultimi anni sono iniziati gli studi sulla capacità degli antibiotici di penetrare nei biofilm di vari microbi.
È stato riscontrato che l'ampicillina penetra scarsamente nei biofilm di Klebsiella pneumoniae e che l'ampicillina, il co-trimaxosolo e la vancomicina penetrano scarsamente nelle comunità di Enterococcus faecalis. L'amoxicillina ampiamente utilizzata penetra male nei biofilm di numerosi microbi.
I fluorochinoloni sono tra gli antibiotici che penetrano bene nei lipidi cellulari. Questo gruppo di farmaci antimicrobici è in grado di agire sui principali agenti patogeni delle malattie urologiche, in concentrazione sufficiente penetra nel fuoco dell'infezione. L'esperienza esistente con l'uso degli antibiotici indica che il processo infettivo, soprattutto le sue manifestazioni cliniche, possono essere affrontati con l'aiuto di antibiotici, sia penetranti che non penetranti nei biofilm. Tuttavia, c'è una differenza tra loro ed è piuttosto significativa. È stato dimostrato che le differenze tra gli antibiotici che penetrano e quelli che non penetrano nei biofilm possono manifestarsi nei risultati a lungo termine del trattamento. L'uso di antibiotici che non penetrano bene nel biofilm porta molto rapidamente alla formazione e selezione di ceppi resistenti. Inoltre, le ricadute si verificano più spesso e si formano focolai di processi cronici.
L'effetto terapeutico sui biofilm può essere mirato ai meccanismi di adesione iniziale dei batteri alla superficie, bloccando la sintesi o la distruzione della matrice polimerica, interrompendo lo scambio di informazioni intercellulari e può anche essere combinato con gli stessi agenti battericidi. Tale trattamento, che influenza la struttura o la funzione dei biofilm, potrebbe essere più efficace della terapia antibiotica standard.
- In contatto con 0
- Google Plus 0
- OK 0
- Facebook 0






