ется секреция соответствующего тройного гормона; при гиперфункции железы секреция соответствующего тропина подавляется. Обратные связи не только позволяют регулировать концентрацию гормонов в крови, но и участвуют в дифференцировке гипоталамуса в онтогенезе. Образование половых гормонов в женском организме происходит циклически, что объ¬ ясняется циклической секрецией гонадотропных гормонов. Синтез этих гормонов контролируется гипоталамусом, образующим рилизинг-фактор этих тропинов (гонадолиберин). Если самке пересадить гипофиз самца, то пересаженный гипофиз начинает функционировать циклично. Половая дифференцировка гипоталамуса происходит под действием андрогенов. Если самца лишить половых желез, продуцирующих андрогены, то гипо¬ таламус будет дифференцироваться по женскому типу.
В железах внутренней секреции иннервированы, как правило, только сосуды, а эндокринные клетки изменяют свою активность лишь под дей¬ ствием метаболитов, кофакторов и гормонов, причем не только гипофизарных. Так, ангиотензин II стимулирует синтез и секрецию альдостерона. Некоторые гормоны гипоталамуса и гипофиза могут образовываться не только в этих тканях. Например, соматостатин обнаружен также в подже¬ лудочной железе, где он подавляет секрецию инсулина и глюкагона.
Большинство нервных и гуморальных путей регуляции сходятся на уровне гипоталамуса, и благодаря этому в организме образуется единая нейроэндокринная регуляторная система. К клеткам гипоталамуса подхо¬ дят аксоны нейронов, расположенных в коре большого мозга и подкорко¬ вых образованиях. Эти аксоны секретируют различные нейромедиаторы, оказывающие на секреторную активность гипоталамуса как активирую¬ щее, так и тормозное влияние. Поступающие из мозга нервные импульсы гипоталамус превращает в эндокринные стимулы, которые могут быть усилены или ослаблены в зависимости от гуморальных сигналов, поступа¬ ющих в гипоталамус от желез и тканей, подчиненных ему.
Тропины, образующиеся в гипофизе, не только регулируют деятель¬ ность подчиненных желез, но и выполняют самостоятельные эндокрин¬ ные функции. Например, пролактин оказывает лактогенное действие, а также тормозит процессы дифференцировки клеток, повышает чувствите¬ льность половых желез к гонадотропинам, стимулирует родительский ин¬ стинкт. Кортикотропин является не только стимулятором стероидогенеза, но и активатором липолиза в жировой ткани, а также важнейшим участ¬ ником процесса превращения в мозге кратковременной памяти в долго¬ временную. Гормон роста может стимулировать активность иммунной си¬ стемы, обмен липидов, Сахаров и др.
В задней доле гипофиза (нейрогипофиз) депонируются антидиуретиче¬ ский гормон (вазопрессин) и окситоцин. Первый вызывает задержку воды в организме и повышает тонус сосудов, второй стимулирует сокращение матки при родах и секрецию молока. Оба гормона синтезируются в гипо¬ таламусе, затем транспортируются по аксонам в заднюю долю гипофиза, где депонируются и потом секретируются в кровь.
Характер процессов, протекающих в ЦНС, во многом определяется со¬ стоянием эндокринной регуляции. Так, андрогены и эстрогены формиру¬ ют половой инстинкт, многие поведенческие реакции. Очевидно, что ней¬ роны, точно так же как и другие клетки нашего организма, находятся под контролем гуморальной системы регуляции. Нервная система, эволюционно более поздняя, имеет как управляющие, так и подчиненные связи с эндокринной системой. Эти две регуляторные системы дополняют друг друга, образуют функционально единый механизм.
4.2. МЕТОДЫ ИССЛЕДОВАНИЯ
Для изучения функций желез внутренней секреции используются экс¬ периментальные и клинические методы исследования. К наиболее важ¬ ным из них следует отнести следующие.
Изучение последствий удаления (экстирпация) эндокринных желез. После удаления какой-либо эндокринной железы возникает комплекс рас¬ стройств, обусловленных выпадением регуляторных эффектов тех гор¬ монов, которые вырабатываются в этой железе. Например, предположе¬ ние о наличии эндокринных функций у поджелудочной железы нашло подтверждение в опытах И. Меринга и О. Минковского (1889), показав¬ ших, что ее удаление у собак приводит к выраженной гипергликемии
и глюкозурии; животные погибали в течение 2-3 нед после операции на фоне явлений тяжелого сахарного диабета. В последующем было установлено, что эти изменения возникают из-за недостатка инсули¬ на - гормона, образующегося в островковом аппарате поджелудочной железы.
Вследствие травматичности оперативного вмешательства вместо хирур¬ гического удаления эндокринной железы может быть использовано введе¬ ние химических веществ, нарушающих их гормональную функцию. На¬ пример, введение животным аллоксана нарушает функцию В-клеток под¬ желудочной железы, что приводит к развитию сахарного диабета, проявле¬ ния которого практически идентичны расстройствам, наблюдаемым после экстирпации поджелудочной железы.
* Наблюдение эффектов, возникших при имплантации желез. Животному с удаленной эндокринной железой можно ее имплантировать заново в хо¬ рошо васкуляризированную область тела, например под капсулу почки или в переднюю камеру глаза. Такая операция называется реимплантацией. Для ее проведения обычно используют эндокринную железу, полу¬ ченную от животного-донора. После реимплантации постепенно восста¬ навливается уровень гормонов в крови, что приводит к исчезновению нарушений, возникших ранее в результате дефицита этих гормонов в ор¬ ганизме. Например, Бертольдом (1849) было показано, что у петухов пе¬ ресадка половых желез в брюшную полость после кастрации предотвра¬ щает развитие посткастрационного синдрома. Возможна также пересад¬ ка эндокринной железы животному, у которого операция экстирпации ранее не производилась. Последнее может быть использовано для изуче¬ ния эффектов, возникающих при избытке гормона в крови, так как его секреция в данном случае осуществляется не только собственной эндок¬ ринной железой животного, но и имплантированной.
А Изучение эффектов, возникших при введении экстрактов эндокринных
желез. Нарушения, возникшие после хирургического удаления эндок¬ ринной железы, могут быть корректированы посредством введения в ор¬ ганизм достаточного количества экстракта данной железы или соответ¬ ствующего гормона.
А Использование радиоактивных изотопов. Иногда для исследования функ¬ циональной активности эндокринной железы может быть использована ее способность извлекать из крови и накапливать определенное соедине¬ ние. Например, щитовидная железа активно поглощает йод, который за¬ тем используется для синтеза тироксина и трийодтиронина. При гипер¬ функции щитовидной железы накопление йода усиливается, при гипо-
функции наблюдается обратное явление. Интенсивность накопления йода может быть определена путем введения в организм радиоактивного изотопа 1 3 1 1 с последующей оценкой радиоактивности щитовидной же¬ лезы. В качестве радиоактивной метки вводят также соединения, кото¬ рые используются для синтеза эндогенных гормонов и включаются в их структуру. В последующем можно определить радиоактивность различ¬ ных органов и тканей и оценить, таким образом, распределение гормона
в организме, а также найти его органы-мишени.
* Определение количественного содержания гормона. В ряде случаев для выяснения механизма какого-либо физиологического эффекта целесо¬ образно сопоставить его динамику с изменением количественного со¬ держания гормона в крови или в другом исследуемом материале.
К наиболее современным относятся методы радиоиммунологического определения концентрации гормонов в крови. Эти методы основаны на том, что меченный радиоактивной меткой гормон и гормон, содержащий¬ ся в исследуемом материале, конкурируют между собой за связывание со специфическими антителами: чем больше в биологическом материале со¬ держится данного гормона, тем меньше свяжется меченых молекул гормо¬ на, так как количество гормонсвязывающих участков в образце постоянно.
* Важное значение для понимания регуляторных функций желез внутрен¬ ней секреции и диагностики эндокринной патологии имеют клинические методы исследования. К ним относятся диагностика типичных симпто¬ мов избытка или недостатка того или иного гормона, использование раз¬ личных функциональных проб, рентгенологические, лабораторные и другие методы исследования.
4.3. ОБРАЗОВАНИЕ, ВЫВЕДЕНИЕ ИЗ ЭНДОКРИННЫХ КЛЕТОК, ТРАНСПОРТ КРОВЬЮ И МЕХАНИЗМЫ ДЕЙСТВИЯ ГОРМОНОВ
4.3.1. Синтез гормонов
В поддержании упорядоченности, согласованности всех физиологиче¬ ских и метаболических процессов в организме участвует более 100 гормо¬ нов и нейромедиаторов. Их химическая природа различна (белки, поли¬ пептиды, пептиды, аминокислоты и их производные, стероиды, производ¬ ные жирных кислот, некоторые нуклеотиды, эфиры и др.). У каждого класса этих веществ пути образования и распада разные.
Белково-пептидные гормоны включают все тропные гормоны, либерины и статины, инсулин, глюкагон, кальцитонин, гастрин, секретин, холецистокинин, ангиотензин II, антидиуретический гормон (вазопрессин), паратгормон и др.
Эти гормоны образуются из белковых предшественников, называемых прогормонами. Как правило, сначала синтезируется препрогормон, из ко¬ торого образуется прогормон, а затем гормон.
Синтез прогормонов осуществляется на мембранах гранулярной эндоплазматической сети (шероховатый ретикулум) эндокринной клетки.
Везикулы с образующимся прогормоном переносятся затем в пластин¬ чатый комплекс Гольджи, где под действием мембранной протеиназы от молекулы прогормона отщепляется определенная часть аминокислотной цепи. В результате образуется гормон, который поступает в везикулы, со-
держащиеся в комплексе Гольджи. В дальнейшем эти везикулы сливаются с плазматической мембраной и высвобождаются во внеклеточное про¬ странство.
Поскольку многие полипептидные гормоны образуются из общего бел¬ кового предшественника, изменение синтеза одного из этих гормонов мо¬ жет приводить к параллельному изменению (ускорение или замедление) синтеза ряда других гормонов. Так, из белка проопиокортина образуются кортикотропин и р-липотропин (схема 4.1), из р-липотропина может обра¬ зоваться еще несколько гормонов: у-липотропин, р-меланоцитостимулиру- ющий гормон, р-эндорфин, у-эндорфин, а-эндорфин, метионин-энкефа- лин.
При действии специфических протеиназ из кортикотропина могут об¬ разовываться а-меланоцитостимулирующий гормон и АКТГ-подобный пептид аденогипофиза. Благодаря сходству структур кортикотропина и а-меланоцитостимулирующего гормона, последний имеет слабую кортикотропную активность. Кортикотропин обладает незначительной способ¬ ностью усиливать пигментацию кожи.
Концентрация белково-пептидных гормонов в крови варьирует в преде¬ лах от 10- 6 до 10- 1 2 М. При стимуляции эндокринной железы концентра¬ ция соответствующего гормона возрастает в 2-5 раз. Так, например, в со¬ стоянии покоя в крови человека содержится около 0,2 мкг АКТГ (в расче¬ те на 5 л крови), а при стрессе это количество возрастает до 0,8-1,0 мкг. В нормальных условиях в крови содержится 0,15 мкг глюкагона и 5 мкг ин¬ сулина. Когда человек голоден, содержание глюкагона может повышаться до 1 мкг, а содержание инсулина снижаться на 40-60 %. После сытного обеда концентрация глюкагона в крови в 1,5-2,8 раза снижается, а содер¬ жание инсулина повышается до 10-25 мкг.
С х е м а 4.1. Образование нескольких белково-пептидных гормонов из одного белкового предшественника под действием стресса
Гипоталамус | |||
Кортиколиберин | |||
Проопиокортин (м.м. 30 000) |
|||
бета -ЛТ(42-134) | |||
гама -ЛТ (42-101) | бета-Эндорфин (104-134) |
||
бета -МСГ(84-101) | мет-Энкефалин (104-108) |
||
Полупериод жизни белково-пептидных гормонов в крови составляет 10-20 мин. Они разрушаются протеиназами клеток-мишеней крови, пече¬ ни, почек.
Стероидные гормоны включают тестостерон, эстрадиол, эстрон, прогес¬ терон, кортизол, альдостерон и др. Эти гормоны образуются из холесте¬ рина в корковом веществе надпочечников (кортикостероиды), а также в семенниках и яичниках (половые стероиды).
В малом количестве половые стероиды могут образовываться в корко¬ вом веществе надпочечников, а кортикостероиды - в половых железах. Свободный холестерин поступает в митохондрии, где превращается в прегненолон, который затем попадает в эндоплазматическую сеть и после этого - в цитоплазму.
В корковом веществе надпочечников синтез стероидных гормонов сти¬ мулируется кортикотропином, а в половых железах - лютеинизирующим гормоном (ЛГ). Эти гормоны ускоряют транспорт эфиров холестерина в эндокринные клетки и активируют митохондриальные ферменты, участву¬ ющие в образовании прегненолона. Кроме того, тропные гормоны активи¬ руют процессы окисления Сахаров и жирных кислот в эндокринных клет¬ ках, что обеспечивает стероидогенез энергией и пластическим материа¬ лом.
Кортикостероиды подразделяют на две группы.Глюкокортикоиды (ти¬ пичный представитель - кортизол) индуцируют синтез ферментов глюконеогенеза в печени, препятствуют поглощению глюкозы мышцами и жировыми клетками, а также способствуют высвобождению из мышц мо¬ лочной кислоты и аминокислот, тем самым ускоряя глюконеогенез в пе¬ чени.
Стимуляция синтеза глюкокортикоидов осуществляется через систему гипоталамус-гипофиз-надпочечники. Стресс (эмоциональное возбужде¬ ние, боль, холод и др.), тироксин, адреналин и инсулин стимулируют вы¬ свобождение кортиколиберина из аксонов гипоталамуса. Этот гормон свя¬ зывается с мембранными рецепторами аденогипофиза и вызывает высво¬ бождение кортикотропина, который с током крови попадает в надпочеч¬ ники и стимулирует там образование глюкокортикоидов - гормонов, по¬ вышающих устойчивость организма к неблагоприятным воздействиям.
Минералокортикоиды (типичный представитель - альдостерон) задер¬ живают натрий в крови. Снижение концентрации натрия в выделяемой моче, а также в секретах слюнных и потовых желез приводит к меньшим потерям воды, так как вода движется через биологические мембраны в на¬ правлении высокой концентрации солей.
Кортикотропин влияет слабо на синтез минералокортикоидов. Имеется дополнительный механизм регуляции синтеза минералокортикоидов, осу¬ ществляющийся через так называемую ренин-ангиотензиновую систему. Рецепторы, реагирующие на давление крови, локализованы в артериолах почек. При снижении давления крови эти рецепторы стимулируют секре¬ цию ренина почками. Ренин является специфической эндопептидазой, от¬ щепляющей от альфа2 -глобулина крови С-концевой декапептид, который на¬ зывают«ангиотензин /». От ангиотензина I карбоксипептидаза (ангиотензинпревращающий фермент, АПФ, расположенный на наружной поверх¬ ности эндотелия кровеносных сосудов) отщепляет два аминокислотных остатка и образует октапептидангиотензин II - гормон, к которому на мембране клеток коркового вещества надпочечников имеются специаль-
ные рецепторы. Связываясь с этими рецепторами, ангиотензин II стиму¬ лирует образование альдостерона, который действует на дистальные кана¬ льцы почек, потовые железы, слизистую оболочку кишечника и увеличи¬ вает в них реабсорбцию ионов Na+ , Cl- и НСО3 - . В результате в крови по¬ вышается концентрация ионов Na+ и снижается концентрация ионов К+ . Эти эффекты альдостерона полностью блокируются ингибиторами синтеза белка.
В крови человека около 500 мкг кортизола. При стрессе его содержание повышается до 2000 мкг. Альдостерона в 1000 раз меньше - около 0,5 мкг. Если человек находится на бессолевой диете, содержание альдостерона повышается до 2 мкг.
Половые стероиды. Андрогены (мужские половые гормоны) продуциру¬ ются интерстициальными клетками яичек и в меньшем количестве яични¬ ками и корковым веществом надпочечников. Основным андрогеном явля¬ ется тестостерон. Этот гормон может претерпевать изменения в клет¬ ке-мишени - превращаться в дигидротестостерон, который обладает бо¬ льшей активностью, чем тестостерон. ЛГ, который стимулирует начальные этапы биосинтеза стероидов в эндокринной железе, активирует также пре¬ вращение тестостерона в дигидротестостерон в клетке-мишени, тем самым усиливая андрогенные эффекты.
Яичники секретируют эстрадиол, андростендион и прогестерон. Фол¬ ликул яичника представляет собой яйцеклетку, окруженную плоскими эпителиальными клетками и соединительнотканной оболочкой. Изнутри эта капсула заполнена фолликулярной жидкостью и зернистыми клетка¬ ми.
При половом созревании синтез этих гормонов начинает контролиро¬ ваться гонадотропинами. При этом фолликулостимулирующий гормон (ФСГ) стимулирует стероидогенез в зернистых клетках, погруженных во внутреннее пространство фолликула, а лютеинизирующий гормон (ЛГ) действует на клетки, формирующие оболочку капсулы. Так как в оболочке образуются мужские половые гормоны (андростерон и тестостерон), а в зернистых клетках они превращаются в женские половые гормоны (эстрон и эстрадиол), очевидно, что для продукции женских половых стероидов должна осуществляться строгая согласованность синтеза и секреции гонадотропинов в гипофизе.
Образование в гипоталамусе гонадолиберина и стимуляция им секре¬ ции ФСГ и ЛГ инициирует механизмы полового созревания. Время начала секреции и количество секретируемого гонадолиберина детерминировано генетически, однако на его секрецию влияют также нейромедиаторы ЦНС: норадреналин, дофамин, серотонин и эндорфины.
Высвобождение гонадолиберина из гипоталамуса обычно происходит во время коротких по своей продолжительности периодов секреции, между которыми наблюдается 2-3-часовая «пауза». Спустя несколько минут после выведения гонадолиберина в крови появляются гонадотропины. Секреция гонадотропинов зависит также от уровня половых стеро¬ идов в крови: эстрогены подавляют выведение ФСГ и стимулируют сек¬ рецию ЛГ гипофизом, а прогестерон тормозит секрецию гонадолиберина в гипоталамусе. Таким образом замыкаются регуляторные связи между сигналами из ЦНС и активностью яичников, осуществляющих стероидо¬ генез.
Ключевую роль в циклическом функционировании женских половых желез играет ФСГ, секреция которого стимулируется гонадолиберином и низким уровнем эстрогенов. ФСГ проводит селекцию только одной из
фолликул (доминантная), которая вступает в менструальный цикл. После этого резко усиливается синтез эстрогенов, что вызывает (по механизму отрицательной обратной связи) снижение уровня ФСГ. Почти одновре¬ менно с этим наблюдается резкий подъем уровня ЛГ, который стимулиру¬ ет созревание доминантной фолликулы, ее разрыв и выход яйцеклетки. Сразу же после этого снижается продукция эстрогенов, что приводит (по механизму отрицательной обратной связи) к подавлению секреции Л Г. Наступает фаза созревания желтого тела, которая сопровождается переме¬ щением яйцеклетки в матку. Это «путешествие» длится 8-9 дней, и, если не происходит оплодотворения яйцеклетки, желтое тело постепенно сни¬ жает продукцию эстрогенов и прогестерона, в результате чего наступает менструация.
Эстрогены (женские половые гормоны) в организме человека в основ¬ ном представлены эстрадиолом. В клетках-мишенях они не метаболизируются.
Действие андрогенов и эстрогенов направлено в основном на органы воспроизведения, проявление вторичных половых признаков, поведенче¬ ские реакции. Андрогенам свойственны также анаболические эффекты - усиление синтеза белка в мышцах, печени, почках. Эстрогены оказывают катаболическое влияние на скелетные мышцы, но стимулируют синтез белка в сердце и печени. Таким образом, основные эффекты половых гор¬ монов опосредуются процессами индукции и репрессии синтеза белка.
Стероидные гормоны легко проникают через клеточную мембрану, по¬ этому их выведение из клетки происходит параллельно с синтезом гормо¬ нов. Содержание стероидов в крови определяется соотношением скоро¬ стей их синтеза и распада. Регуляция этого содержания осуществляется главным образом путем изменения скорости синтеза. Тропные гормоны (кортикотропин, ЛГ и ангиотензин) стимулируют этот синтез. Устранение тропного влияния приводит к торможению синтеза стероидных гормонов.
Действующие концентрации стероидных гормонов составляют 10- 1 1 -10- 9 М. Период их полураспада равен 1/2-1 1/2 ч.
Тиреоидные гормоны включают тироксин и трийодтиронин. Синтез этих гормонов осуществляется в щитовидной железе, в которой ионы йода окисляются при участии пероксидазы до йодиниум-иона, способно¬ го йодировать тиреоглобулин - тетрамерный белок, содержащий около 120 тирозинов. Йодирование тирозиновых остатков происходит при учас¬ тии пероксида водорода и завершается образованием монойодтирозинов и дийодтирозинов. После этого происходит «сшивка» двух йодированных тирозинов. Эта окислительная реакция протекает с участием пероксида¬ зы и завершается образованием в составе тиреоглобулина трийодтиронина и тироксина. Для того чтобы эти гормоны освободились из связи с белком, должен произойти протеолиз тиреоглобулина. При расщеплении одной молекулы этого белка образуется 2-5 молекул тироксина (Т4 ) и трийодтиронина (Т3 ), которые секретируются в молярных соотношениях, равных 4:1.
Синтез и выведение тиреоидных гормонов из продуцирующих их кле¬ ток находятся под контролем гипоталамо-гипофизарной системы. Тиреотропин активирует аденилатциклазу щитовидной железы, ускоряет актив-
ный транспорт йода, а также стимулирует рост эпителиальных клеток щи¬ товидной железы. Эти клетки формируют фолликул, в полости которого происходит йодирование тирозина. Адреналин и простагландин Е2 также могут повышать концентрацию цАМФ в щитовидной железе, при этом они вызывают такое же стимулирующее влияние на синтез тироксина, как и тиреотропин.
Активный транспорт ионов йода в железу при действии тиреотропина происходит против 500-кратного градиента. Тиреотропин стимулирует также синтез рибосомальной РНК и мРНК тиреоглобулина, т.е. происхо¬ дит усиление как транскрипции, так и трансляции белка, служащего ис¬ точником тирозинов для синтеза Т3 и Т4 . Выведение Т3 и Т4 из клеток - их продуцентов - осуществляется посредством пиноцитоза. Частички коллоида окружаются мембраной эпителиальной клетки и поступают в цитоплазму в виде пиноцитозных пузырьков. При слиянии этих пузырь¬ ков с лизосомами эпителиальной клетки происходит расщепление тиреог¬ лобулина, который составляет основную массу коллоида, что приводит к выделению Т3 и Т4 . Тиреотропин и другие факторы, повышающие концен¬ трацию цАМФ в щитовидной железе, стимулируют пиноцитоз коллоида, процесс образования и движения секреторных пузырьков. Таким образом, тиреотропин ускоряет не только синтез, но и выведение Т3 и Т4 из клетокпродуцентов. При повышении уровня Т3 и Т4 в крови подавляется секре¬ ция тиреолиберина и тиреотропина.
Тиреоидные гормоны могут циркулировать в крови в неизменном виде в течение нескольких дней. Такая устойчивость гормонов объясняется, по-видимому, образованием прочной связи с Т4 -связывающими глобули¬ нами и преальбуминами в плазме крови. Эти белки имеют в 10-100 раз большее сродство к Т4 , чем к Т3 , поэтому в крови человека содержится 300-500 мкг Т4 и лишь 6-12 мкг Т3 .
Катехоламины включают адреналин, норадреналин и дофамин. Источ¬ ником катехоламинов, как и тиреоидных гормонов, служит тирозин. Кате¬ холамины, образующиеся в мозговом веществе надпочечников, выделяют¬ ся в кровь, а не в синаптическую щель, т.е. являются типичными гормона¬ ми.
В некоторых клетках синтез катехоламинов заканчивается образовани¬ ем дофамина, а адреналин и норадреналин образуются в меньшем количе¬ стве. Такие клетки есть в составе гипоталамуса.
Синтез катехоламинов в мозговом веществе надпочечников стимулиру¬ ется нервными импульсами, поступающими по чревному симпатическому нерву. Выделяющийся в синапсах ацетилхолин взаимодействует с холинергическими рецепторами никотинового типа и возбуждает нейросекреторную клетку надпочечника. Благодаря существованию нервно-рефлек¬ торных связей надпочечники отвечают усилением синтеза и выделения ка¬ техоламинов в ответ на болевые и эмоциональные раздражители, гипок¬ сию, мышечную нагрузку, охлаждение и др. Подобный тип регуляции эн¬ докринной железы, являющийся исключением из обычного правила, мож¬ но объяснить тем, что мозговой слой надпочечника в эмбриогенезе обра¬ зуется из нервной ткани, поэтому у него сохраняется типичный нейрональный тип регуляции. Существуют и гуморальные пути регуляции актив¬ ности клеток мозгового вещества надпочечников: синтез и выделение ка¬ техоламинов могут возрастать под действием инсулина, глюкокортикоидов при гипогликемии.
Катехоламины подавляют как собственный синтез, так и выделение. В адренергических синапсах на пресинаптической мембране есть адьфа-адре-
нергические рецепторы. При выбросе катехоламинов в синапс эти рецеп¬ торы активируются и оказывают ингибирующее влияние на секрецию ка¬ техоламинов.
Гематоэнцефалический барьер не пропускает катехоламины из крови в мозг. В то же время диоксифенилаланин, их предшественник, легко про¬ никает через этот барьер и может усилить образование катехоламинов в мозге.
Катехоламины инактивируются в тканях-мишенях, печени и почках. Решающее значение в этом процессе играют два фермента - моноаминоксидаза, расположенная на внутренней мембране митохондрий, и кате- хол-О-метилтрансфераза, цитозольный фермент.
Эйкозаноиды включают простагландины, тромбоксаны и лейкотриены. Эйкозаноиды называют гормоноподобными веществами, так как они мо¬ гут оказывать только местное действие, сохраняясь в крови в течение не¬ скольких секунд. Образуются во всех органах и тканях практически всеми типами клеток.
Биосинтез большинства эйкозаноидов начинается с отщепления арахидоновой кислоты от мембранного фосфолипида или диацилглицерина в плазматической мембране. Синтетазный комплекс представляет собой полиферментную систему, функционирующую преимущественно на мем¬ бранах эндоплазматической сети. Образующиеся эйкозаноиды легко про¬ никают через плазматическую мембрану клетки, а затем через межкле¬ точное пространство переносятся на соседние клетки и выходят в кровь и лимфу. Наиболее интенсивно простагландины образуются в яичках и яичниках.
Простагландины могут активировать аденилатциклазу, тромбоксаны увеличивают активность фосфоинозитидного обмена, а лейкотриены по¬ вышают проницаемость мембран для Са2 + . Поскольку цАМФ и Са2 + сти¬ мулируют синтез эйкозаноидов, замыкается положительная обратная связь в синтезе этих специфических регуляторов.
Период полураспада эйкозаноидов составляет 1-20 с. Ферменты, инактивирующие их, имеются практически во всех тканях, но наибольшее их количество содержится в легких.
4.3.2. Выведение гормонов из клеток-продуцентов и транспорт гормонов кровью
Стероидные гормоны благодаря своей липофильности не накапливаются в эндокринных клетках, а легко проходят через мембрану и поступают в кровь и лимфу. В связи с этим регуляция содержания этих гормонов в крови осуществляется путем изменения скорости их синтеза.
Тиреоидные гормоны также липофильны и также легко проходят через мембрану, однако они ковалентно связаны в эндокринной железе с тиреоглобулином, поэтому могут выводиться из клетки только после наруше¬ ния этой связи. Чем больше йодированных тирозилов в составе тиреоглобулина и чем выше скорость протеолиза йодированного белка, тем больше тиреоидных гормонов в крови. Регуляция содержания тиреоидных гормо¬ нов осуществляется двумя путями - ускорением как процессов йодирова¬ ния, так и разрушения тиреоглобулина.
Гормоны, имеющие белковую и пептидную природу, а также катехолами¬ ны, гистамин, серотонини др. - это гидрофильные вещества, которые не могут диффундировать через клеточную мембрану. Для выведения этих
молекул созданы специальные механизмы, чаще всего пространственно и функционально разобщенные с процессами биосинтеза.
Многие белково-пептидные гормоны образуются из предшественников большой молекулярной массы, и выведение этих гормонов становится возможным только после того, как произойдет отщепление «лишнего» фрагмента. Так, выведению инсулина из клетки предшествует превраще¬ ние в В-клетках поджелудочной железы препроинсулина в проинсулин, а затем в инсулин. Биосинтез инсулина и других белково-пептидных гормо¬ нов, а также их транспорт к периферии секреторной клетки занимает обычно 1-3 ч. Очевидно, что воздействие на биосинтез приведет к изме¬ нению уровня белкового гормона в крови лишь через несколько часов. Влияние же на выведение этих гормонов, синтезированных «впрок» и за¬ пасенных в специальных везикулах, позволяет повышать их концентрацию в несколько раз за секунды или минуты.
Для секреции белково-пептидных гормонов и катехоламинов необходи¬ мы ионы Са2 + . Принято считать, что для выведения гормонов важна не собственно деполяризация мембраны, а происходящий при ней вход Са2 + в цитоплазму клетки.
Поступив в кровь, гормоны связываются с транспортными белками, что защищает их от разрушения и экскреции. В связанной форме гормон с током крови переносится от места секреции к клеткам-мишеням. В этих клетках есть рецепторы, которые имеют большее сродство к гормону, чем белки крови.
Обычно лишь 5-10 % молекул гормона находится в крови в свободном состоянии, и только свободные молекулы могут взаимодействовать с ре¬ цептором. Однако, как только они свяжутся с рецептором, равновесие в реакции взаимодействия гормона с транспортными белками сдвигается в сторону распада комплекса и концентрация свободных молекул гормона останется практически неизменной. При избытке гормонсвязывающих белков в крови концентрация свободных молекул гормона может снизить¬ ся до критической величины.
Связывание гормонов в крови зависит от их сродства к связывающим белкам и концентрации этих белков. К их числу относятся транскортин, связывающий кортикостероиды, тестостерон-эстрогенсвязывающий гло¬ булин, тироксинсвязывающий глобулин, тироксинсвязывающий преальбумин и др. Едва ли не все гормоны могут связываться с альбумином, кон¬ центрация которого в крови в 1000 раз больше, чем концентрация других гормонсвязывающих белков. Однако сродство к альбумину у гормонов в десятки тысяч раз меньше, поэтому с альбуминами обычно связано 5- 10% гормонов, а со специфическими белками 85-90 %. Альдостерон, по-видимому, не имеет специфических «транспортных» белков, поэтому находится преимущественно в связи с альбумином.
4.3.3. Молекулярные механизмы действия гормонов
Гормоны, действующие через мембранные рецепторы и системы вто¬ ричных посредников, стимулируют химическую модификацию белков. Наиболее хорошо изучено фосфорилирование. Регуляция, происходящая за счет химических процессов (синтез и расщепление вторичного посред¬ ника, фосфорилирование и дефосфорилирование белка), развивается и га¬ сится за минуты или десятки минут.
115. Основные системы межклеточной коммуникации: эндокринная, паракринная, аутокринная регуляция.
По расстоянию от клетки-продуцента гормона до клетки-мишени различают эндокринный, паракринный и аутокринный варианты регуляции.
Эндокринная
, или дистантная, регуляция. Секреция гормона происходит в жидкие среды организма. Клетки-мишени могут отстоять от эндокринной клетки сколь угодно далеко. Пример: секреторные клетки эндокринных желёз, гормоны из которых поступают в систему общего кровотока.
Паракринная регуляция
. Продуцент биологически активного вещества и клетка-мишень расположены рядом. Молекулы гормона достигают мишени путём диффузии в межклеточном веществе. Например, в париетальных клетках желёз желудка секрецию Н + стимулируют гастрин и гистамин, а подавляют соматостатин и Пг, секретируемые рядом расположенными клетками.
Аутокринная регуляция
. При аутокринной регуляции клетка-продуцент гормона имеет рецепторы к этому же гормону (другими словами, клетка-продуцент гормона в то же время является его мишенью). Примеры: эндотелины, вырабатываемые клетками эндотелия и воздействующие на эти же эндотелиальные клетки; Т-лимфоциты, секретирующие интерлейкины, имеющие мишенями разные клетки, в том числе и Т-лимфоциты.
116. Роль гормонов в системе регуляции метаболизма. Клетки-мишени и клеточные рецепторы гормонов
Роль гормонов в регуляции обмена веществ и функций . Интегрирующими регуляторами, связывающими различные регуляторные механизмы и метаболизм в разных органах, являются гормоны. Они функционируют как химические посредники, переносящие сигналы, возникающие в различных органах и ЦНС. Ответная реакция клетки на действие гормона очень разнообразна и определяется как химическим строением гормона, так и типом клетки, на которую направлено действие гормона. В крови гормоны присутствуют в очень низкой концентрации. Для того чтобы передавать сигналы в клетки, гормоны должны распознаваться и связываться особыми белками клетки - рецепторами, обладающими высокой специфичностью. Физиологический эффект гормона определяется разными факторами, например концентрацией гормона (которая определяется скоростью инактивации в результате распада гормонов, протекающего в основном в печени, и скоростью выведения гормонов и его метаболитов из организма), его сродством к белкам-переносчикам (стероидные и тиреоидные гормоны транспортируются по кровеносному руслу В комплексе с белками), количеством и типом рецепторов на поверхности клеток-мишеней. Синтез и секреция гормонов стимулируются внешними и внутренними сигналами, поступающими в ЦНС.Эти сигналы по нейронам поступают в гипоталамус, где стимулируют синтез пептидных рилизинг-гормонов (от англ,release - освобождать) - либеринов и статинов, которые, соответственно, стимулируют или ингибируют синтез и секрецию гормонов передней доли гипофиза. Гормоны передней доли гипофиза, называемые тройными гормонами, стимулируют образование и секрецию гормонов периферических эндокринных желёз, которые поступают в общий кровоток и взаимодействуют с клетками-мишенями. Поддержание уровня гормонов в организме обеспечивает механизм отрицательной обратной связи. Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные железы, либо на гипоталамус. Синтез и секреция тропных гормонов подавляется гормонами эндокринных периферических желёз. Такие петли обратной связи действуют в системах регуляции гормонов надпочечников, щитовидной железы, половых желёз. Не все эндокринные железы регулируются подобным образом. Гормоны задней доли гипофиза (вазопрессин и окситоцин) синтезируются в гипоталамусе в виде предшественников и хранятся в гранулах терминальных аксонов нейрогипофиза. Секреция гормонов поджелудочной железы (инсулина и глюкагона) напрямую зависит от концентрации глюкозы в крови. В регуляции межклеточных взаимодействий участвуют также низкомолекулярные белковые соединения - цитокины. Влияние цитокинов на различные функции клеток обусловлено их взаимодействием с мембранными рецепторами. Через образование внутриклеточных посредников сигналы передаются в ядро, где происходят активация определённых генов и индукция синтеза белков. Все цитокины объединяются следующими общими свойствами:
- синтезируются в процессе иммунного ответа организма, служат медиаторами иммунной и воспалительной реакций и обладают в основном аутокринной, в некоторых случаях паракринной и эндокринной активностью;
- действуют как факторы роста и факторы дифференцировки клеток (при этом вызывают преимущественно медленные клеточные реакции, требующие синтеза новых белков);
- обладают плейотропной (полифункциональной) активностью.
Биологическое действие гормонов проявляется через их взаимодействие с рецепторами клеток-мишеней. Для проявления биологической активности связывание гормона с рецептором должно приводить к образованию химического сигнала внутри клетки, который вызывает специфический биологический ответ, например изменение скорости синтеза ферментов и других белков или изменение их активности. Мишенью для гормона могут служить клетки одной или нескольких тканей. Воздействуя на клетку-мишень, гормон вызывает специфическую ответную реакцию. Например, щитовидная железа - специфическая мишень для тиреотропина, под действием которого увеличивается количество ацинарных клеток щитовидной железы, повышается скорость биосинтеза тиреоидных гормонов. Глюкагон, воздействуя на адипоциты, активирует липолиз, в печени стимулирует мобилизацию гликогена и глюконеогенез. Характерный признак клетки-мишени - способность воспринимать информацию, закодированную в химической структуре гормона.
Рецепторы гормонов . Начальный этап в действии гормона на клетку-мишень - взаимодействие гормона с рецептором клетки. Концентрация гормонов во внеклеточной жидкости очень низка и обычно колеблется в пределах 10 -6 -10 -11 ммоль/л. Клетки-мишени отличают соответствующий гормон от множества других молекул и гормонов благодаря наличию на клетке-мишени соответствующего рецептора со специфическим центром связывания с гормоном.
Общая характеристика рецепторов
Рецепторы пептидных гормонов и адреналина располагаются на поверхности клеточной мембраны. Рецепторы стероидных и тиреоидных гормонов находятся внутри клетки. Причём внутриклеточные рецепторы для одних гормонов, например глюкокортикоидов, локализованы в цитозоле, для других, таких как андрогены, эстрогены, тиреоидные гормоны, расположены в ядре клетки. Рецепторы по своей химической природе являются белками и, как правило, состоят из нескольких доменов. В структуре мембранных рецепторов можно выделить 3 функционально разных участка. Первый домен (домен узнавания) расположен в N-концевой части полипептидной цепи на внешней стороне клеточной мембраны; он содержит гликозилированные участки и обеспечивает узнавание и связывание гормона. Второй домен - трансмембранный. У рецепторов одного типа, сопряжённых с G-белками, он состоит из 7 плотно упакованных α-спиральных полипептидных последовательностей. У рецепторов другого типа трансмембранный домен включает только одну α-спирадизованную полипептидную цепь (например, обе β-субъединицы гетеротетрамерного рецептора инсулина α 2 β 2). Третий (цитоплазматический) домен создаёт химический сигнал в клетке, который сопрягает узнавание и связывание гормона с определённым внутриклеточным ответом. Цитоплазматический участок рецептора таких гормонов, как инсулин, фактор роста эпидермиса и инсулиноподобный фактор роста-1 на внутренней стороне мембраны обладает тирозинки-назной активностью, а цитоплазматические участки рецепторов гормона роста, пролактина и цитокинов сами не проявляют тирозинкиназ-ную активность, а ассоциируются с другими цитоплазматическими протеинкиназами, которые их фосфорилируют и активируют.
Рецепторы стероидных и тиреоидных гормонов содержат 3 функциональные области. На С-концевом участке полипептидной цепи рецептора находится домен узнавания и связывания гормона. Центральная часть рецептора включает домен связывания ДНК. На N-концевом участке полипептидной цепи располагается домен, называемый вариабельной областью рецептора, отвечающий за связывание с другими белками, вместе с которыми участвует в регуляции транскрипции.
117. Механизмы передачи гормональных сигналов в клетки.
По механизму действия гормоны можно разделить на 2 группы. К первой группе относят гормоны, взаимодействующие с мембранными рецепторами (пептидные гормоны, адреналин, а также гормоны местного действия - цитокины, эйкозаноиды). Вторая группа включает гормоны, взаимодействующие с внутриклеточными рецепторами.Связывание гормона (первичного посредника) с рецептором приводит к изменению кон-формации рецептора. Это изменение улавливается другими макромолекулами, т.е. связывание гормона с рецептором приводит к сопряжению одних молекул с другими (трансдукция сигнала). Таким образом, генерируется сигнал, который регулирует клеточный ответ путём изменения активности или количества ферментов и других белков. В зависимости от способа передачи гормонального сигнала в клетках меняется скорость реакций метаболизма:
- в результате изменения активности ферментов;
- в результате изменения количества ферментов
118. Классификация гормонов по химическому строению и биологическим функциям
Классификация гормонов по химическому строению
| Пептидные гормоны | Стероиды | Производные аминокислот |
| Адренокортикотропный гормон (кортикотропин, АКТГ) | Альдостерон | Адреналин |
| Гормон роста (соматотропин, ГР, СТГ) | Кортизол | Норадреналин |
| Тиреотропный гормон (тиреотропин, ТТГ) | Кальцитриол | Трийодтиронин (Т 3) |
| Лактогенный гормон (пролактин, ЛТГ) | Тестостерон | Тироксин (Т 4) |
| Лютеинизирующий гормон (лютропин, ЛГ) | Эстрадиол | |
| Фолликулостимулирующий гормон (ФСГ) | Прогестерон | |
| Меланоцитстимулирующий гормон (МСГ) | ||
| Хорионический гонадотропин (ХГ) | ||
| Антидиуретический гормон (вазопрессин, АДГ) | ||
| Окситоцин | ||
| Паратиреоидный гормон (паратгормон, ПТГ) | ||
| Кальцитонин | ||
| Инсулин | ||
| Глюкагон |
Классификация гормонов по биологическим функциям*
| Регулируемые процессы | Гормоны |
| Обмен углеводов, липйдов, аминокислот | Инсулин, глюкагон, адреналин, кортизол, тироксин, соматотропин |
| Водно-солевой обмен | Альдостерон, антидиуретический гормон |
| Обмен кальция и фосфатов | Паратгормон, кальцитонин, кальцитриол |
| Репродуктивная функция | Эстрадиол, тестостерон, прогестерон, гонадотропные гормоны |
| Синтез и секреция гормонов эндокринных желёз | Тропные гормоны гипофиза, либерины и статины гипоталамуса |
| Изменение метаболизма в клетках, синтезирующих гормон | Эйкозаноиды, гистамин, секретин, гастрин, соматостатин, вазоактивный интестинальный пептид (ВИП), цитокины |
(*) Эта классификация условна, поскольку одни и те же гормоны могут выполнять разные функции
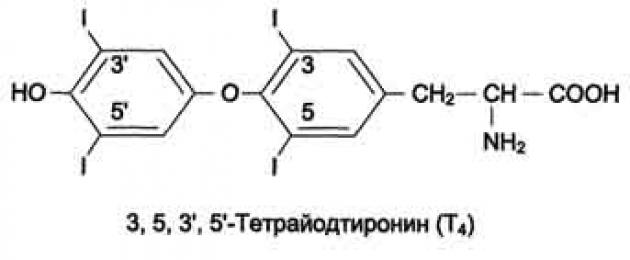
119. Строение, синтез и метаболизм иодтиронинов. Влияние на обмен веществ. Изменение метаболизма при гипо- и гипертиреозе. Причины и проявление эндемического зоба.
Биосинтез йодтиронинов . Йодтиронины синтезируются в составе белка тиреоглобулина (Тг) в фолликулах, которые представляют собой морфологическую и функциональную единицу щитовидной железы.
Тиреоглобулин - гликопротеин с молекулярной массой 660 кД, содержащий 115 остатков тирозина. 8-10% массы тиреоглобулина представлено углеводами. Содержание йодида в организме составляет 0,2-1%
. 
Тиреоглобулин синтезируется на рибосомах шероховатого ЭР в виде претиреоглобулина, затем переносится в цистерны ЭР, где происходит формирование вторичной и третичной структуры, включая процессы гликозилирования. Из цистерн ЭР Тиреоглобулин поступает в аппарат Гольджи, включается в состав секреторных гранул и секретируется во внеклеточный коллоид, где происходит йодирование остатков тирозина и образование йодтиронинов. Йодирование тиреоглобулина и образование йодтиронинов осуществляется в несколько этапов
Транспорт йода в клетки щитовидной железы . Йод в виде органических и неорганических соединений поступает в ЖКТ с пищей и питьевой водой. Суточная потребность в йоде составляет 150-200 мкг. 25-30% этого количества йодидов захватывается щитовидной железой. Транспорт йодида в клетки щитовидной железы - энергозависимый процесс и происходит при участии специального транспортного белка против электрохимического градиента (соотношение концентраций I - в железе к концентрации I - в сыворотке крови в норме составляет 25:1). Работа этого йодид-переносящего белка сопряжена с Nа + ,К + -АТФ-азой.
Окисление йода. Окисление I - в I + происходит при участии гемсодержащей тиреоперокси-дазы и Н 2 О 2 в качестве окислителя. Йодирование тирозина . Окисленный йод взаимодействует с остатками тирозина в молекуле тиреоглобулина. Эта реакция также катализируется тиреопероксидазой.
Образование йодтиронинов. Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием монойод-тирозинов (МИТ) и дийодтирозинов (ДИТ). Две молекулы ДИТ конденсируются с образованием йодтиронина Т 4 , а МИТ и ДИТ - с образованием йодтиронина Т 3 . Йодтиреоглобулин транспортируется из коллоида в фолликулярную клетку путём эндоцитоза и гидролизуется ферментами лизосом с освобождением Т 3 и Т 4 . В нормальных условиях щитовидная железа сек-ретирует 80-100 мкг Т 4 и 5 мкг Т 3 в сутки. Ещё 22-25 мкг Т 3 образуется в результате дейодирования Т 4 в периферических тканях по 5"-углеродному атому.
Транспорт и метаболизм йодтиронинов . От половины до двух третей Т 3 и Т 4 находятся в организме вне щитовидной железы. Большая часть их циркулирует в крови в связанной форме в комплексе с белками: тироксинсвязывающим глобулином (ТСГ) и тироксинсвязывающим преальбумином (ТСПА). ТСГ служит основным транспортным белком йодтиронинов, а также формой их депонирования. Он обладает более высоким сродством к Т 3 и Т 4 и в нормальных условиях связывает почти всё количество этих гормонов. Только 0,03% Т 4 и 0,3% Т 3 находятся в крови в свободной форме. Т 1/2 Т 4 в плазме в 4-5 раз больше, чем Т 3 . Для Т 4 этот период составляет около 7 дней, а для Т 3 - 1-1,5 дня. Биологическая активность йодтиронинов обусловлена несвязанной фракцией. Т 3 - основная биологически активная форма йодтиронинов; его сродство к рецептору клеток-мишеней в 10 раз выше, чем у Т 4 . В периферических тканях в результате дейодирования части Т 4 по пятому углеродному атому образуется так называемая "реверсивная" форма Т 3 , которая почти полностью лишена биологической активности. Другие пути метаболизма йодтиронинов включают полное дейодирование, дезаминирование или декарбоксилирование. Йодированные продукты катаболизма йодтиронинов конъюгируют-ся в печени с глюкуроновой или серной кислотами, секретируются с жёлчью, в кишечнике вновь всасываются, дейодируются в почках и выделяются с мочой.
Механизм действия и биологические функции йодтиронинов. Клетки-мишени йодтиронинов имеют 2 типа рецепторов к этим гормонам. Основные эффекты йодтиронинов - результат их взаимодействия с высокоспецифичными рецепторами, которые в комплексе с гормонами постоянно находятся в ядре и взаимодействуют с определёнными последовательностями ДНК, участвуя в регуляции экспрессии генов. Другие рецепторы расположены в плазматической мембране клеток, но это не те же самые белки, что в ядре. Они обладают более низким сродством к йодтиронинам и, вероятно, обеспечивают связывание гормонов для удержания их в непосредственной близости к клетке. При физиологической концентрации йодтиронинов их действие проявляется в ускорении белкового синтеза, стимуляции процессов роста и клеточной дифференцировки. В этом отношении йодтиронины - синергисты гормона роста. Кроме того, Т 3 ускоряет транскрипцию гена гормона роста. У животных при дефиците Т 3 клетки гипофиза теряют способность к синтезу гормона роста. Очень высокие концентрации Т 3 тормозят синтез белков и стимулируют катаболические процессы, показателем чего служит отрицательный азотистый баланс. Метаболические эффекты йодтиронинов относят в основном к энергетическому метаболизму, что проявляется в повышении поглощения клетками кислорода. Этот эффект проявляется во всех органах, кроме мозга, РЭС и гонад. В разных клетках Т 3 стимулирует работу Nа + ,К + -АТФ-азы, на что затрачивается значительная часть энергии, утилизируемой клеткой. В печени йодтиронины ускоряют гликолиз, синтез холестерола и синтез жёлчных кислот. В печени и жировой ткани Т 3 повышает чувствительность клеток к действию адреналина и косвенно стимулирует липолиз в жировой ткани и мобилизацию гликогена в печени. В физиологических концентрациях Т 3 увеличивает в мышцах потребление глюкозы, стимулирует синтез белков и увеличение мышечной массы, повышает чувствительность мышечных клеток к действию адреналина. Йодтиронины также участвуют в формировании ответной реакции на охлаждение увеличением теплопродукции, повышая чувствительность симпатической нервной системы к норадреналину и стимулируя секрецию норадреналина.
Заболевания щитовидной железы Гормоны щитовидной железы необходимы для нормального развития человека.
Гипотиреоз у новорождённых приводит к развитию кретинизма, который проявляется множественными врождёнными нарушениями и тяжёлой необратимой задержкой умственного развития. Гипотиреоз развивается вследствие недостаточности йодтиронинов. Обычно гипотиреоз связан с недостаточностью функции щитовидной железы, но может возникать и при заболеваниях гипофиза и гипоталамуса.
Наиболее тяжёлые формы гипотиреоза, сопровождающиеся слизистым отёком кожи и подкожной клетчатки, обозначают термином "микседема " (от греч. туха - слизь, oedema - отёк). Отёчность обусловлена избыточным накоплением гликозаминогликанов и воды. В подкожной клетчатке накапливается глюкуроновая и в меньшей степени хондроитинсерная кислоты. Избыток гликозаминогликанов вызывает изменения коллоидной структуры межклеточного матрикса, усиливает его гидрофильность и связывает ионы натрия, что приводит к задержке воды. Характерные проявления заболевания: снижение частоты сердечных сокращений, вялость, сонливость, непереносимость холода, сухость кожи. Эти симптомы развиваются вследствие снижения основного обмена, скорости гликолиза, мобилизации гликогена и жиров, потребления глюкозы мышцами, уменьшения мышечной массы и снижения теплопродукции. При возникновении гипотиреоза у детей старшего возраста наблюдают отставание в росте без задержки умственного развития. В настоящее время у взрослых людей частой причиной гипотиреоза является хронический аутоиммунный тиреоидит, приводящий к нарушению синтеза йодтиронинов (зоб Хашимото ).
Гипотиреоз может быть также результатом недостаточного поступления йода в организм -эндемический зоб . Эндемический зоб (нетоксический зоб) часто встречается у людей, живущих в районах, где содержание йода в воде и почве недостаточно. Если поступление йода в организм снижается (ниже 100 мкг/сут), то уменьшается продукция йодтиронинов, что приводит к усилению секреции ТТГ (из-за ослабления действия йодтиронинов на гипофиз по механизму отрицательной обратной связи), под влиянием которого происходит компенсаторное увеличение размеров щитовидной железы (гиперплазия), но продукция йодтиронинов при этом не увеличивается.
Гипертиреоз возникает вследствие повышенной продукции йодтиронинов. Диффузный токсический зоб (базедова болезнь, болезнь Грейвса) - наиболее распространённое заболевание щитовидной железы. При этом заболевании отмечают увеличение размеров щитовидной железы (зоб), повышение концентрации йодтиронинов в 2-5 раз и развитие тиреотоксикоза. Характерные признаки тиреотоксикоза: увеличение основного обмена, учащение сердцебиений, мышечная слабость, снижение массы тела (несмотря на повышенный аппетит) , потливость, повышение температуры тела, тремор и экзофтальм (пучеглазие). Эти симптомы отражают одновременную стимуляцию йодтиронинами как анаболических (рост и дифференцировка тканей), так и катаболических (катаболизм углеводов, ли-пидов и белков) процессов. В большей мере усиливаются процессы катаболизма, о чём свидетельствует отрицательный азотистый баланс. Гипертиреоз может возникать в результате различных причин: развитие опухоли, тиреоидит, избыточное поступление йода и йодсодер-жащих препаратов, аутоиммунные реакции. Болезнь Грейвса возникает в результате образования антител к тиреоидным антигенам. Один из них, иммуноглобулин (IgG), имитирует действие тиреотропина, взаимодействуя с рецепторами тиреотропина на мембране клеток щитовидной железы. Это приводит к диффузному разрастанию щитовидной железы и избыточной неконтролируемой продукции Т 3 и Т 4 , поскольку образование IgG не регулируется по механизму обратной связи. Уровень ТТГ при этом заболевании снижен вследствие подавления функции гипофиза высокими концентрациями йодтиронинов.
120. Регуляция энергетического метаболизма, роль инсулина и контринсулярных гормонов в обеспечении гомеостаза .
Основные пищевые вещества (углеводы, жиры, белки) окисляются в организме с освобождением свободной энергии, которая используется в анаболических процессах и при осуществлении физиологических функций. Энергетическая ценность основных пищевых веществ выражается в килокалориях и составляет: для углеводов - 4 ккал/г, для жиров - 9 ккал/г, для белков - 4 ккал/г. Взрослому здоровому человеку в сутки требуется 2000-3000 ккал (8000-12 000 кДж) энергии. При обычном ритме питания промежутки между приёмами пищи составляют 4-5 ч с 8-12-часовым ночным перерывом. Во время пищеварения и абсорбтивного периода (2-4 ч) основные энергоносители, используемые тканями (глюкоза, жирные кислоты, аминокислоты), могут поступать непосредственно из пищеварительного тракта. В постабсорбтивном периоде и при голодании энергетические субстраты образуются в процессе катаболизма депонированных энергоносителей. Изменения в потреблении энергоносителей и энергетических затратах координируются путём чёткой регуляции метаболических процессов в разных органах и системах организма, обеспечивающей энергетический гомеостаз. Основную роль в поддержании энергетического гомеостаза играют гормоны инсулин и глюкагон , а также другие контринсулярные гормоны - адреналин, кортизол, йодтиронины и соматотропин. Инсулин и глюкагон играют главную роль в регуляции метаболизма при смене абсорбтивного и постабсорбтивного периодов и при голодании. Абсорбтивный период характеризуется временным повышением концентрации глюкозы, аминокислот и жиров в плазме крови. Клетки поджелудочной железы отвечают на это повышение усилением секреции инсулина и снижением секреции глюкагона. Увеличение отношения инсулин/глюкагон вызывает ускорение использования метаболитов для запасания энергоносителей: происходит синтез гликогена, жиров и белков. Режим запасания включается после приёма пищи и сменяется режимом мобилизации запасов после завершения пищеварения. Тип метаболитов, которые потребляются, депонируются и экспортируются, зависит от типа ткани. Главные органы, связанные с изменениями потока метаболитов при смене режимов мобилизации и запасания энергоносителей, - печень, жировая ткань и мышцы.
Изменения метаболизма в печени в абсорбтивном периоде
После приёма пищи печень становится главным потребителем глюкозы, поступающей из пищеварительного тракта. Почти 60 из каждых 100 г глюкозы, транспортируемой портальной системой, задерживается в печени. Увеличение потребления печенью глюкозы - не результат ускорения её транспорта в клетки (транспорт глюкозы в клетки печени не стимулируется инсулином), а следствие ускорения метаболических путей, в которых глюкоза превращается в депонируемые формы энергоносителей: гликоген и жиры. При повышении концентрации глюкозы в гепатоцитах происходит активация глюкокиназы, превращающей глюкозу в глюкозо-6-фосфат. Глюкокиназа имеет высокое значение К m для глюкозы, что обеспечивает высокую скорость фосфорилирования при высоких концентрациях глюкозы. Кроме того, глюкокиназа не ингибируется глюкозо-6-фосфатом (см. раздел 7). Инсулин индуцирует синтез мРНК глюкокиназы. Повышение концентрации глюкозо-6-фосфата в гепатоцитах обусловливает ускорение синтеза гликогена. Этому способствуют одновременная инактивация гликогенфосфорилазы и активация гликогенсинтазы. Под влиянием инсулина в гепатоцитах ускоряется гликолиз в результате повышения активности и количества ключевых ферментов: глюкокиназы, фосфофруктокиназы и пируваткиназы. В то же время происходит торможение глюконеогенеза в результате инактивации фруктозо-1,6-бисфосфатазы и снижения количества фосфоенолпируваткарбоксикиназы - ключевых ферментов глюконеогенеза. Повышение концентрации глюкозо-6-фосфата в гепатоцитах в абсорбтивном периоде, сочетается с активным использованием NADPH для синтеза жирных кислот, что способствует стимуляции пентозофосфатного пути. Ускорение синтеза жирных кислот обеспечивается доступностью субстратов (ацетил-КоА и NADPH), образующихся при метаболизме глюкозы, а также активацией и индукцией ключевых ферментов синтеза жирных кислот. В абсорбтивном периоде в печени ускоряется синтез белков. Однако количество аминокислот, поступающих в печень из пищеварительного тракта, превышает возможности их использования для синтеза белков и других азотсодержащих соединений. Излишек аминокислот либо поступает в кровь и транспортируется в другие ткани, либо дезаминируется с последующим включением безазотистых остатков в общий путь катаболизма.
Изменения метаболизма в адипоцитах . Основная функция жировой ткани - запасание энергоносителей в форме триацилгли-церолов. Под влиянием инсулина ускоряется транспорт глюкозы в адипоциты. Повышение внутриклеточной концентрации глюкозы и активация ключевых ферментов гликолиза обеспечивают образование ацетил-КоА и глицерол-3-фосфата, необходимых для синтеза ТАГ. Стимуляция пентозофосфатного пути обеспечивает образование NADPH, необходимого для синтеза жирных кислот. Однако биосинтез жирных кислот de novo в жировой ткани человека протекает с высокой скоростью только после предшествующего голодания. При нормальном ритме питания для синтеза ТАГ используются в основном жирные кислоты, поступающие из ХМ и ЛПОНП под действием ЛП-липазы. Вместе с тем при увеличении отношения инсулин/глюкагон гормончувствительная ТАГ-липаза находится в дефосфорилированной неактивной форме, и процесс липолиза тормозится.
Изменение метаболизма в мышцах в абсорбтивном периоде . В абсорбтивном периоде под влиянием инсулина ускоряется транспорт глюкозы в клетки мышечной ткани. Глюкоза фосфорилируется и окисляется для обеспечения клетки энергией, а также используется для синтеза гликогена. Жирные кислоты, поступающие из ХМ и ЛПОНП, в этот период играют незначительную роль в энергетическом обмене мышц. Поток аминокислот в мышцы и биосинтез белков также увеличиваются под влиянием инсулина, особенно после приёма белковой пищи.
Эндокринная система регулирует множество функций разных клеток и органов. Эта регуляторная функция осуществляется при помощи сигнальных молекул - гормонов, вырабатываемых эндокринными клетками, циркулирующих во внутренней среде организма и связывающихся со специфическими рецепторами гормонов на соответствующих клетках-мишенях.
Химия гормонов. По химическому строению различают следующие типы гормонов: олигопептид (например, нейропептиды); полипептид (например, инсулин); гликопротеин (например, тиреотропин); стероид (например, альдостерон и кортизол); производное тирозина (например, йодсодержащие гормоны щитовидной железы: трийодтиронин - Т 3 и тироксин - T 4); эйкозаноиды (метаболиты арахидоновой кислоты).
Цитология эндокринных клеток. Эндокринные клетки имеют строение, определяемое химической природой синтезируемого гормона.
Пептиды, белки, гликопротеины, катехоловые амины. Для этих эндокринных клеток характерно наличие гранулярной эндоплазматической сети (здесь происходит сборка пептидной цепи), комплекса Гольджи (присоединение углеводных остатков, формирование секреторных гранул), секреторных гранул.
Стероидные гормоны. Для клеток, синтезирующих стероидные гормоны, характерно присутствие развитой гладкой эндоплазматической сети и многочисленных митохондрий.
Тропный гормон - гормон, клетками-мишенями которого являются другие эндокринные клетки (например, часть эндокринных клеток передней доли гипофиза синтезирует и секретирует в кровь АКТГ (адренокортикотропный гормон). Мишени АКТГ - эндокринные клетки пучковой зоны коры надпочечников, синтезирующие глюкокортикоиды.
Рилизинг-гормоны (рилизинг-факторы) [от англ. releasing hormone (releasing factor)] - группа синтезируемых в нейронах гипоталамической области мозга гормонов, мишенями которых являются эндокринные клетки передней доли гипофиза (например, рилизинг-гормон для синтезирующих АКТГ клеток передней доли гипофиза - кортиколиберин). Рилизинг-гормоны подразделяют на либерины и статины.
Рис. 9-5. Варианты воздействия гормонов-лигандов на клетки-мишени.
Либерин - рилизинг-гормон, способствующий усилению синтеза и секреции соответствующего гормона в эндокринных клетках передней доли гипофиза.
Статин - рилизинг-гормон, в отличие от либеринов ингибирующий синтез и секрецию гормонов в клетках-мишенях.
Варианты эндокринной регуляции. В зависимости от расстояния от продуцента гормона до клетки-мишени различают эндокринный, паракринный и аутокринный варианты регуляции (рис. 9-5).
Эндокринная, или дистантная регуляция. Секреция гормона происходит во внутреннюю среду, клетки-мишени могут отстоять от эндокринной клетки сколь угодно далеко. Наиболее яркий пример: секреторные клетки эндокринных желёз, гормоны из которых поступают в систему общего кровотока.
Паракринная регуляция. Продуцент биологически активного вещества и клеткамишень расположены рядом, молекулы гормона достигают мишени путём диффузии в межклеточном веществе. Например, в париетальных клетках желёз желудка секрецию H + стимулируют гастрин и гистамин, а подавляют соматостатин и простагландины, секретируемые рядом расположенными клетками.
Аутокринная регуляция. При аутокринной регуляции сама клетка-продуцент гормона имеет рецепторы к этому же гормону (другими словами, клетка-продуцент гормона в то же время является собственной мишенью). В качестве примера приведём эндотелины, вырабатываемые клетками эндотелия и воздействующие на эти же эндотелиальные клетки.
Классификация. Органы эндокринной системы подразделяют на несколько групп:
гипоталамо-гипофизарная система: нейросекреторные нейроны и аденогипофиз;
мозговые придатки: гипофиз и эпифиз;
бранхиогенная группа (имеющая происхождение из эпителия глоточных карманов): щитовидная железа, паращитовидные железы, вилочковая железа;
надпочечниково-адреналовая система: кора надпочечников, мозговое вещество надпочечников и параганглии;
островки поджелудочной железы;
диффузная эндокринная система: эндокринные клетки, рассеянные в различных органах.
ГИПОТАЛАМО-ГИПОФИЗАРНАЯ СИСТЕМА
Эпителиального генеза передняя доля гипофиза (синтез тропных гормонов, экспрессия гена проопиомеланокортина), перикарионы нейросекреторных нейронов гипоталамуса (синтез рилизинг-гормонов, вазопрессина, окситоцина, орексинов), гипоталамо-гипофизарный тракт (транспорт гормонов по аксонам нейросекреторных нейронов), аксо-вазальные синапсы (секреция вазопрессина и окситоцина в капилляры задней доли гипофиза, секреция рилизинг-гормонов в капилляры срединного возвышения), портальная система кровотока между срединным возвышением и передней долей гипофиза в совокупности формируют гипоталамо-гипофизарную систему (рис. 9-6, рис. 9-12).
Гипофиз
Гипофиз анатомически состоит из ножки и тела, а гистологически подразделяется на адено- и нейрогипофиз.
Развитие гипофиза. Гипофиз образуется из двух зачатков эктодермального (карман Ратке) и нейрогенного (processus infundibularis).
Карман Ратке. На 4-5-й неделе эктодермальный эпителий крыши ротовой бухты образует карман Ратке - вырост, направляющийся к мозгу. Из этого гипофизарного кармана развивается аденогипофиз (передняя, промежуточная и входящая в состав ножки гипофиза туберальная доли).
Processus infundibularis. Навстречу карману Ратке растёт выпячивание промежуточного мозга, дающее начало нейрогипофизу (задняя доля гипофиза, нейрогипофизарная часть ножки гипофиза и отчасти срединное возвышение).
Кровоснабжение гипофиза. Портальная система кровотока состоит из первичной капиллярной сети срединного возвышения, воротных вен туберальной части аденогипофиза и вторичной капиллярной сети передней доли (рис. 9-9). Приносящие гипофизарные артерии в медиобазальном гипоталамусе (срединное возвышение) образуют первичную капиллярную сеть. Терминали аксонов нейросекреторных клеток
 Рис. 9-6. Анатомия гипофиза.
Рис. 9-6. Анатомия гипофиза.
гипоталамуса заканчиваются на этих капиллярах. Кровь из первичной капиллярной сети собирается в портальные вены, идущие по гипофизарной ножке (туберальная часть) в переднюю долю. Здесь портальные вены переходят в капилляры вторичной сети. Обогащённая гормонами передней доли кровь из вторичной капиллярной сети поступает в общую циркуляцию через выносящие вены.
Аденогипофиз (см. рис. 9-6) состоит из передней и промежуточной долей и туберальной части ножки гипофиза. Аденогипофиз покрыт фиброзной капсулой. Передняя доля представлена тяжами эндокринных клеток (аденоцитов), окружённых сетью ретикулиновых волокон. В передней доле ретикулиновые волокна окружают капилляры с фенестрированным эндотелием и широким просветом (синусоиды) вторичной капиллярной сети. Туберальная часть состоит из тяжей эпителиальных клеток, между ними расположены гипофизарные воротные вены (vv. portae hypophysis, см. рис. 9-9), соединяющие первичную капиллярную сеть (срединное возвышение) и вторичную капиллярную сеть (передняя доля гипофиза). Эндокринная функция эпителиальных клеток туберальной части отсутствует, в ней изредка встречаются базофильные аденоциты. Средняя (промежуточная) доля гипофиза у человека выражена слабо.
Аденогипофиз
 Рис. 9-9. Система кровоснабжения гипофиза.
ПЕРЕДНЯЯ ДОЛЯ
Рис. 9-9. Система кровоснабжения гипофиза.
ПЕРЕДНЯЯ ДОЛЯ
Передняя доля - эпителиальная эндокринная железа, её клетки синтезируют и секретируют тропные гормоны и продукты экспрессии гена проопиомеланокортина. Разные эндокринные клетки передней доли синтезируют различные пептидные гормоны. Эндокринные клетки передней доли содержат элементы гранулярной эндоплазматической сети, комплекс Гольджи, многочисленные митохондрии и секреторные гранулы различного диаметра. Клетки расположены анастомозирующими тяжами и островками между кровеносными капиллярами с фенестрированным эндотелием. В последние выводятся гормоны, а из капилляров к клеткам поступают либерины и статины.
Классификация эндокринных клеток передней доли (аденоцитов) основана на связывании стандартных красителей, по этому признаку различают хромофильные (базофильные и оксифильные) и хромофобные (плохо окрашивающиеся) клетки. Хромофобные клетки - гетерогенная популяция, включающая дегранулировавшие клетки (оксифилы и базофилы разных типов) и камбиальный резерв. Регенерация аденоцитов происходит из клеток камбиального резерва.
Базофильные аденоциты подразделяют на кортикотрофы, тиротрофы и гонадотрофы.
♦ Кортикотрофы экспрессируют ген проопиомеланокортина и содержат гранулы диаметром около 200 нм.
♦ Тиротрофы синтезируют тиреотрофный гормон (ТТГ) и содержат мелкие (около 150 нм) гранулы.
♦ Гонадотрофы синтезируют фолликулостимулирующий гормон (фоллитропин) и лютропин, размеры гранул варьируют от 200 до 400 нм. Фоллитропин и лютропин синтезируются в разных подтипах гонадотрофов.
Ацидофильные аденоциты синтезируют, накапливают в гранулах и секретируют соматотрофин (гормон роста) и пролактин.
♦ Соматотрофы имеют гранулы диаметром до 400 нм.
♦ Лактотрофы содержат мелкие (около 200 нм) гранулы. При беременности и лактации величина гранул может достигать 600 нм.
В передней доле синтезируются СТГ (соматотрофный гормон, соматотро[ф][п]ин, гормон роста), ТТГ (тиреотропный гормон, тиротрофин), АКТГ (адренокортикотропный гормон), гонадотропины (гонадотропные гормоны), а именно лютеинизирующий гормон (лютропин) и фолликулостимулирующий гормон (фоллитропин), а также пролактин. Экспрессия гена проопиомеланокортина приводит к синтезу и секреции ряда пептидов (АКТГ, β- и γ-липотропины, α-, β- и γ-меланотропины, β-эндорфин), из которых гормональная функция установлена для АКТГ и меланотропинов; функции остальных пептидов изучены недостаточно.
Гормоны роста
К этой группе относят гипофизарный гормон роста и хорионический соматомаммотрофин.
♦ Гормон роста гипофизарный (СТГ, соматотрофин, соматотрофный гормон) нормально экспрессируется только в ацидофильных клетках (соматотрофах) передней доли гипофиза.
♦ Хорионическийсоматомаммотрофин синтезируетсяв клеткахсинцитиотрофобласта. Этот гормон известен также как плацентарный лактоген.
Нативный гормон роста - полипептидная цепь, состоящая из 191 аминокислотных остатков. Синтез и секрецию СТГ стимулирует соматолиберин, а подавляет соматостатин. Эффекты гормона роста
опосредуют соматомедины (инсулиноподобные факторы роста, IGF), синтезируемые преимущественно в гепатоцитах. СТГ - анаболический гормон, стимулирующий рост всех тканей. Наиболее очевидны эффекты СТГ на рост длинных трубчатых костей.
Меланокортины и АКТГ
Адренокортикотропный гормон, α-, β- и γ-меланоцитостимулирующие гормоны (меланотропины), липотропины и β-эндорфин образуются из молекулы-предшественника - проопиомеланокортина (POMC). Продукты гена POMC все вместе называют меланокортинами. Адренокортикотропный гормон. АКТГ состоит из 39 аминокислот. Синтез АКТГ осуществляют кортикотрофы преимущественно передней и в меньшей степени промежуточной доли гипофиза, а также некоторые нейроны ЦНС. Гипоталамический кортиколиберин стимулирует синтез и секрецию АКТГ, а АКТГ стимулирует синтез и секрецию гормонов коры надпочечников (главным образом, глюкокортикоидов).
Гонадотропные гормоны
В эту группу входят гипофизарные фоллитропин и лютропин, а также хорионический гонадотропин (ХГТ) плаценты. Гонадотропные гормоны, а также тиротропин (ТТГ) - гликопротеины, состоящие из двух субъединиц (СЕ). Структура α-СЕ фоллитропина, лютропина, ХГТ и ТТГ идентична, а структура β-СЕ тех же гормонов различна. Гипоталамический гонадолиберин стимулирует синтез и секрецию фоллитропина и лютропина в базофилах (гонадотрофы) передней доли гипофиза. Фоллитропин (фолликулостимулирующий гормон). α-Ингибин - пептидный гормон, вырабатываемый зернистыми клетками фолликулов яичника и сустентоцитами яичка, - подавляет секрецию фоллитропина. Фоллитропин, как и лютропин, регулирует овариальный цикл у женщин. У мужчин мишени фоллитропина - сустентоциты яичка (регуляция сперматогенеза).
Лютропин (лютеинизирующий гормон). У женщин лютропин, как и фоллитропин, регулирует овариальный цикл и эндокринную функцию яичников. У мужчин лютропин стимулирует синтез тестостерона в интерстициальных эндокриноцитах яичек.
Хорионический гонадотропин (ХГТ) - гликопротеин, синтезируемый клетками трофобласта с 10-12 дней развития. При беременности ХГТ взаимодействует с клетками жёлтого тела (синтезирующего и секретирующего прогестерон) яичников.
Тиреотропный гормон
Тиротропин (тиреотропный гормон, ТТГ) синтезируется в базофильных клетках (тиротрофы) передней доли гипофиза. Соматостатин по-
давляет секрецию ТТГ, а гипоталамический тиролиберин стимулирует синтез и секрецию ТТГ. Гормоны щитовидной железы (Т 3 и Т 4), циркулирующие в крови, регулируют секрецию ТТГ по принципу отрицательной обратной связи. Увеличение содержания свободных T 4 и T 3 подавляет секрецию ТТГ. Уменьшение содержания свободных T 4 и T 3 стимулирует секрецию тиротропина. Рецептор ТТГ экспрессируется в фолликулярных клетках щитовидной железы, а также в ретробульбарных тканях. Тиротропин стимулирует дифференцировку эпителиальных клеток щитовидной железы (кроме т.н. светлых клеток, синтезирующих тирокальцитонин) и их функциональное состояние (включая синтез тироглобулина и секрецию Т 3 и Т 4).
Пролактин
Синтез пролактина происходит в ацидофильных аденоцитах (лактотрофах) передней доли гипофиза. Количество лактотрофов составляет не менее трети всех эндокринных клеток аденогипофиза. При беременности объём передней доли удваивается за счёт увеличения числа лактотрофов и их гипертрофии. Пролактиностатин подавляет секрецию пролактина из лактотрофов. Дофамин ингибирует синтез и секрецию пролактина. Тиролиберин стимулирует секрецию пролактина из лактотрофов. Стимуляция соска и околососкового поля увеличивает секрецию пролактина. Главная функция пролактина - регулирование функции молочной железы.
Нейрогипофиз
Нейрогипофиз (задняя доля гипофиза и нейрогипофизарная часть ножки гипофиза) состоит из клеток нейроглии - питуицитов и кровеносных сосудов. Собственная эндокринная функция питуицитов неизвестна, но нейрогипофиз содержит аксоны гипоталамо-гипофизарного тракта и их окончания на кровеносных капиллярах (аксо-вазальные синапсы). Эти аксоны принадлежат нейронам, расположенным в паравентрикулярном и супраоптическом ядрах гипоталамуса (рис. 9-12). Большие нейроны этих ядер продуцируют вазопрессин и окситоцин, которые по аксонам транспортируются в заднюю долю, где и происходит их высвобождение из нейросекреторных клеток. Следовательно, задняя доля, как и передняя, служит местом выделения пептидных гормонов из гипоталамуса.
Аксо-вазальные синапсы образованы терминальными расширениями аксонов нейросекреторных нейронов гипоталамуса, контактирующими со стенкой кровеносных капилляров срединного возвышения и задней доли гипофиза. Аксоны имеют локальные утолщения (нейросекреторные тельца), заполненные пузырьками и гранулами с гормонами.
Гипоталамус
Нейросекреторные нейроны гипоталамуса - типичные нервные клетки. В перикарионах этих нейронов синтезируются рилизинг-гормоны, орексины, АДГ, окситоцин и другие гормоны. Такие гормон-продуцирующие нервные клетки входят в состав многих ядер гипоталамуса, в т.ч. надзрительного (n. supraopticus) и околожелудочкового (n. paraventricularis).
Гипоталамо-гипофизарный тракт образован аксонами нейросекреторных нейронов гипоталамуса (рис. 9-12). Синтезируемые в нейросекреторных нейронах гормоны при помощи аксонного транспорта достигают аксо-вазальных синапсов нейрогипофиза.
Гипоталамические рилизинг-гормоны
В нейросекреторных нейронах гипоталамуса синтезируются либерины [гонадолиберин (люлиберин), кортиколиберин, соматолиберин, ти- ролиберин] и статины (меланостатин, пролактиностатин, соматостатин).
Соматостатин синтезируется многими нейронами ЦНС, δ-клетками панкреатических островков, эндокринными клетками пищеварительного тракта и ряда других внутренних органов. Соматостатин - мощный регулятор функций эндокринной и нервной систем, ингибирует синтез и секрецию множества гормонов и секретов.
Кортистатин продуцируется ГАМКергическими нейронами коры большого мозга и гиппокампа. Этот пептид связывается с рецепторами соматостатина и обладает общими с соматостатином свойствами.
Соматолиберин стимулирует секрецию гормона роста в передней доле гипофиза.
Гонадолиберин и пролактиностатин. Ген LHRH кодирует структуру гонадолиберина и пролактиностатина. Мишени гонадолиберина - гонадотрофы, а пролактиностатина - лактотрофы передней доли гипофиза. Гонадолиберин - ключевой нейрорегулятор репродуктивной функции, стимулирует синтез и секрецию фоллитропина и лютропина в продуцирующих гонадотрофы клетках, а пролактиностатин подавляет секрецию пролактина из лактотрофных клеток передней доли гипофиза.
Тиролиберин синтезируется многими нейронами ЦНС (в т.ч. нейросекреторными нейронами околожелудочкового ядра). Мишени тиролиберина - тиротрофы и лактотрофы передней доли гипофиза. Тиролиберин стимулирует секрецию пролактина из лактотрофов и секрецию тиротропина из тиротрофов.
Кортиколиберин синтезируется в нейросекреторных нейронах околожелудочкового ядра гипоталамуса, некоторых других нейронах ЦНС, а также в эндометрии, плаценте, матке, яичнике, яичках, желудке, кишечнике, надпочечниках, щитовидной железе и в коже. Кортиколиберин стимулирует синтез АКТГ и других продуктов экспрессии гена проопиомеланокортина (POMC) клетками аденогипофиза. Кортиколиберин, продуцируемый в матке и плаценте, может играть важную роль в нормальном течении беременности.
Меланостатин подавляет образование меланотропинов.

Рис. 9-12. Гипоталамо-гипофизарн ^1 й тракт. Нейроны с перикарионами больших размеров, локализованные в гипоталамусе, секретируют рилизинг-гормоны в просвет капилляров в области срединного возвышения и воронки, где расположены капилляры первичной сети, собирающие кровь в длинные портальные вены. По ним гипоталамические рилизинг-гормоны поступают в ножку гипофиза и далее в капилляры передней доли (вторичная капиллярная сеть). Аксоны малых нейросекреторных клеток спускаются в ножку гипофиза и выделяют рилизинг-гормоны в капиллярное сплетение, расположенное непосредственно в ножке. Короткие портальные вены переносят рилизинг-гормоны во вторичную капиллярную сеть передней доли. Большие нейроны паравентрикулярного и супраоптического ядер гипоталамуса синтезируют вазопрессин и окситоцин. По аксонам этих нейросекреторных клеток данные гормоны поступают в заднюю долю, где выделяются из нервных терминалей и поступают в просвет многочисленных сосудов, образующих здесь сплетение.
Орексины
В латеральном гипоталамусе расположены нейросекреторные нервные клетки, синтезирующие орексины (гипокретины) A и B. Орексины функционируют как регуляторы сна и бодрствования, участвуют в регуляции пищевого поведения.
Гормоны задней доли
Гормоны задней доли - аргинин вазопрессин (антидиуретический гормон, АДГ), окситоцин, а также нейрофизины - синтезируются в нейросекреторных нейронах надзрительного и околожелудочкового ядер гипоталамуса. Содержащие гормоны мембранные пузырьки транспортируются по аксонам этих нейронов в составе гипоталамогипофизарного тракта в заднюю долю гипофиза и через аксо-вазальные синапсы гормоны секретируются в кровь.
Окситоцин - циклический нонапептид. Мишени окситоцина - ГМК миометрия и миоэпителиальные клетки молочной железы. Окситоцин стимулирует сокращение ГМК миометрия в родах, при оргазме, в менструальную фазу. Окситоцин стимулирует продукцию и секрецию пролактина, секретируется при раздражении соска и околососкового поля, стимулирует сокращение миоэпителиальных клеток альвеол лактирующей молочной железы (рефлекс молокоотделения). Окситоцин регулирует поведенческую активность, связанную с беременностью и родами.
Аргинин вазопрессин - нонапетид. Экспрессия АДГ происходит в части нейросекреторных нейронов околожелудочкового и надзрительного ядер гипоталамуса. Секрецию АДГ стимулирует через барорецепторы каротидной области гиповолемия, т.е. уменьшение объёма циркулирующей крови, а ингибируют алкоголь, α-адренергические агонисты, глюкокортикоиды. Аргинин вазопрессин оказывает антидиуретический (регулятор реабсорбции воды в собирательных трубочках почки) и сосудосуживающий (вазоконстриктор) эффекты. Главная функция АДГ - регуляция обмена воды (поддержание постоянного осмотического давления жидких сред организма).
Нейрофизины I и II кодируются генами окситоцина и АДГ соответственно. Нейрофизины относят к связывающим окситоцин и АДГ белкам.
ЭПИФИЗ
Шишковидная железа - небольшой (5-8 мм) конической формы вырост промежуточного мозга, соединённый ножкой со стенкой третьего желудочка. Капсула органа образована соединительной тканью мягкой мозговой оболочки. От капсулы отходят перегородки, содержащие кровеносные сосуды и сплетения симпатических нервных волокон. Эти перегородки частично подразделяют тело железы на дольки. Паренхима органа состоит из пинеалоцитов и интерстициальных (глиальных) клеток. В интерстиции присутствуют отложения солей кальция, известные как «мозговой песок» (corpora arenacea). Иннервация: орган снабжён многочисленными постганглионарными нервными волокнами от верхнего шейного симпатического узла. Функция органа у человека изучена слабо, хотя железа у ряда позвоночных выполняет различные функции [например, у некоторых амфибий и рептилий эпифиз содержит фоторецепторные элементы (т.н. теменной глаз)], иногда бездоказательно переносимые на человека. Эпифиз у человека,
скорее всего, - звено реализации биологических ритмов, в т.ч. околосуточных.
Пинеалоциты содержат крупное ядро, хорошо развитую гладкую эндоплазматическую сеть, элементы гранулярной эндоплазматической сети, свободные рибосомы, комплекс Гольджи, множество секреторных гранул, микротрубочки и микрофиламенты. Многочисленные длинные отростки пинеалоцитов заканчиваются расширениями на капиллярах и среди клеток эпендимы. Пинеалоциты синтезируют гормон мелатонин и серотонин.
Мелатонин (N-ацетил-5-метокситриптамин) секретируется в цереброспинальную жидкость и в кровь преимущественно в ночные часы.
Серотонин (5-гидрокситриптамин) синтезируется преимущественно в дневные часы. Интерстициальные клетки напоминают астроциты, имеют многочисленные ветвящиеся отростки, округлое плотное ядро, элементы гранулярной эндоплазматической сети и структуры цитоскелета: микротрубочки, промежуточные филаменты и множество микрофиламентов. Циркадианный ритм, или околосуточный ритм - один из биологических ритмов (суточная, помесячная, сезонная и годовая ритмика), скоординированный с суточной цикличностью вращения Земли; несколько не соответствует 24 часам. Многие процессы, в т.ч. гипоталамическая нейросекреция, подчиняются околосуточному ритму. Механизмы околосуточного ритма. Изменения освещённости через зрительный тракт оказывают влияние на разряды нейронов надперекрестного ядра (nucleus suprachiasmaticus) ростро-вентральной части гипоталамуса. Надзрительное ядро содержит т.н. эндогенные часы - неизвестной природы генератор биологических ритмов (включая околосуточный), контролирующий продолжительность сна и бодрствования, пищевое поведение, секрецию гормонов и т.д. Сигнал генератора - гуморальный фактор, секретируемый из надзрительного ядра (в т.ч. в цереброспинальную жидкость). Сигналы от надзрительного ядра через нейроны околожелудочкового ядра (n. paraventricularis) активируют преганглионарные симпатические нейроны боковых столбов спинного мозга. Симпатические преганглионары активируют нейроны верхнего шейного узла. Постганглионарные симпатические волокна от верхнего шейного узла секретируют норадреналин, взаимодействующий с α- и β-адренорецепторами плазмолеммы пинеалоцитов. Активация адренорецепторов приводит к увеличению внутриклеточного содержания цАМФ и экспрессии гена CREM, а также к транскрипции арилалкиламин-N-ацетилтрансферазы, фермента синтеза мелатонина.
ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа секретирует регуляторы основного обмена - йодсодержащие гормоны - трийодтиронин (Т 3) и тироксин (Т 4), а также кальцитонин, один из эндокринных регуляторов обмена Ca 2+ . Йодсодержащие гормоны вырабатывают эпителиальные клетки стенки фолликулов, кальцитонин - светлые клетки.
Развитие. Эпителий бранхиогенной группы желёз (щитовидная, вилочковая, околощитовидные) развивается из энтодермы глоточных карманов. В конце 3-го месяца развития плода начинается синтез йодсодержащих гормонов, появляющихся в амниотической жидкости. Синтезирующие кальцитонин светлые клетки (С-клетки) щитовидной железы развиваются из нервного гребня.
ПАРЕНХИМА
Паренхима щитовидной железы - совокупность секретирующих тиреоидные гормоны клеток и С-клеток, синтезирующих кальцитонин. И те, и другие входят в состав фолликулов и скоплений межфолликулярных клеток.
Тиреоциты и йодсодержащие гормоны
Фолликулы - различной величины и формы (преимущественно округлые) пузырьки, содержащие коллоид. Стенка фолликула образована эпителиальными фолликулярными клетками (продукция йодсодержащих гормонов), прикреплёнными к базальной мембране. Между базальной мембраной и фолликулярными клетками встречаются более крупные светлые клетки (синтез кальцитонина). Фолликулярные клетки, или тироциты образуют стенку фолликула и формируют его содержимое, синтезируя и секретируя в коллоид тироглобулин. Фермент тиропероксидаза и рецептор N-ацетилглюко- замина также синтезируются в фолликулярных клетках. Основная функция фолликулярных клеток - синтез и секреция T 4 и T 3 - складывается из многих процессов: образование тироглобулина → секреция тироглобулина в полость фолликула → поглощение йода из крови - окисление йода - йодирование тироглобулина в полости фолликула → эндоцитоз и расщепление тироглобулина → секреция T 3 и T 4 . Функцию фолликулярных клеток стимулирует тиротропин (ТТГ). Форма клеток (от низкой кубической до цилиндрической) эпителиальной стенки фолликула зависит от интенсивности их функционирования: высота клеток пропорциональна напряжённости осуществляемых в них процессов.
Базальная часть клеток содержит ядро, гладкий и шероховатый эндоплазматический ретикулум. В плазмолемму встроены связанные с G-белком рецепторы ТТГ, Na + /I - -котранспортёр. Возможна складчатость плазмолеммы (отражает интенсивность обмена между клетками и кровеносными капиллярами - захват йода, поступление метаболитов, секреция гормонов).
Латеральная часть клеток содержит межклеточные контакты, предупреждающие просачивание коллоида.
Апикальная часть содержит выраженный комплекс Гольджи (формирование секреторных пузырьков, присоединение углеводов к тироглобулину), разные типы пузырьков [секреторные (содержат тироглобулин), окаймлённые (незрелый тироглобулин из полости фолликула поступает в клетку для рециклизации и выве-
дения в кровоток), эндоцитозные (содержат зрелый тироглобулин для его последующей деградации в фаголизосомах)], микроворсинки (увеличение поверхности обмена между клетками и полостью фолликула). Апикальная плазмолемма содержит рецепторы N-ацетилгалактозамина (связывание незрелого тироглобулина для его интернализации путём опосредованного этими рецепторами эндоцитоза), рецепторы мегалина (интернализация, трансцитоз и секреция в кровь тироглобулина), анионообменники (перемещение йода из цитоплазмы клетки в полость фолликула). В связи с мембранными структурами апикальной части клеток находится тиропероксидаза. Продукция йодсодержащих гормонов. Синтез и секреция йодсодержащих гормонов включает несколько этапов (рис. 9-17). Йодсодержащие гормоны. Тироксин (T 4) и трийодтиронин (T 3) - водонерастворимые соединения, поэтому сразу после секреции в кровь гормоны образуют комплексы с транспортными белками плазмы, которые не только обеспечивают циркуляцию Т 3 и Т 4 в крови, но и предотвращают деградацию и экскрецию этих гормонов.
Тироксин (3,5,3",5"-тетрайодтиронин, C 15 H 11 I 4 NO 4 , M r 776,87) - основной йодсодержащий гормон, на долю T 4 приходится не менее 90% секретируемых щитовидной железой йодсодержащих гормонов.
♦ L-форма тироксина физиологически примерно вдвое активнее рацемической (DL-тироксин), D-форма гормональной активности не имеет.
♦ Дейодирование наружного кольца тироксина приводит к образованию Т 3 .
♦ Дейодирование внутреннего кольца тироксина приводит к образованию реверсивного Т 3 (rT 3), имеющего незначительную физиологическую активность.
Трийодотиронин (3,5,3"-трийодтиронин, C 15 H 12 I 3 NO 4 , M r 650,98). На долю T 3 приходится лишь 10% содержащихся в крови йодсодержащих гормонов, но физиологическая активность T 3 примерно в четыре раза выше, чем тироксина.
Функции йодсодержащих гормонов многочисленны. Например, Т 3 и Т 4 увеличивают обменные процессы, ускоряют катаболизм белков, жиров и углеводов, эти гормоны необходимы для нормального развития ЦНС, они стимулируют рост хряща и поддерживают рост кости, увеличивают частоту сердечных сокращений и сердечный выброс. Крайне разнообразные эффекты йодсодержащих гормонов на клетки-мишени (ими практически являются все клетки организма) объясняют увеличением синтеза белков и потребления кислорода.
С-клетки
С-клетки в составе фолликулов называют также парафолликулярными клетками. В них происходит экспрессия кальцитонинового гена CALC1, кодирующего кальцитонин, катакальцин и относящийся к кальцитониновому гену пептид α. С-клетки крупнее тироцитов, в составе фолликулов расположены, как правило, одиночно. Морфология этих клеток характерна для клеток, синтезирующих белок на экспорт (присутствуют шероховатая эндоплазматическая сеть, комплекс Голь-
 Рис. 9-17. Биосинтез йодсодержащих гормонов. 1.
Йод поступает в тироцит через Na + /I - -котранспортёр. 2.
Из цитоплазмы в полость фолликула йодид транспортируется через анионообменник SAT. 3.
На границе апикальной мембраны тироцита и коллоида тиропероксидаза катализирует окисление йодида с образованием молекулы йода. 4.
Тиропероксидаза
катализирует йодирование остатков тирозина в молекуле тироглобулина с
образованием монойодтирозина и дийодтирозина. 5.
Синтез трийодтиронина и тетрайодтиронина. 6.
Интернализация йодированного тироглобулина путём эндоцитоза. 7.
Слияние эндоцитозного пузырька с лизосомой и деградация тироглобулина. 8.
Высвобождение монойодтирозина, дийодтирозина, Т3 и Т4 в цитоплазму клетки. 9.
Дейодирование и реутилизация монойодтирозина и дийодтирозина. 10.
Секреция йодсодержащих гормонов в кровь.
Рис. 9-17. Биосинтез йодсодержащих гормонов. 1.
Йод поступает в тироцит через Na + /I - -котранспортёр. 2.
Из цитоплазмы в полость фолликула йодид транспортируется через анионообменник SAT. 3.
На границе апикальной мембраны тироцита и коллоида тиропероксидаза катализирует окисление йодида с образованием молекулы йода. 4.
Тиропероксидаза
катализирует йодирование остатков тирозина в молекуле тироглобулина с
образованием монойодтирозина и дийодтирозина. 5.
Синтез трийодтиронина и тетрайодтиронина. 6.
Интернализация йодированного тироглобулина путём эндоцитоза. 7.
Слияние эндоцитозного пузырька с лизосомой и деградация тироглобулина. 8.
Высвобождение монойодтирозина, дийодтирозина, Т3 и Т4 в цитоплазму клетки. 9.
Дейодирование и реутилизация монойодтирозина и дийодтирозина. 10.
Секреция йодсодержащих гормонов в кровь.
джи, секреторные гранулы, митохондрии). На гистологических препаратах цитоплазма С-клеток выглядит светлее цитоплазмы тироцитов, отсюда их название - светлые (clear) клетки.
Кальцит онин - пептид, содержащий 32 аминокислотных остатка.
♦ Регулятор экспрессии - Са 2+ плазмы крови, внутривенное его введение существенно увеличивает секрецию кальцитонина.
♦ Функции кальцитонина, как одного из регуляторов кальциевого обмена, определяют как антагонистические функциям гормона паращитовидной железы.
Катакальцин - пептид, состоящий из 21 аминокислотного остатка, имеет те же функции, что и кальцитонин.
Относящиеся к кальцитониновому гену пептиды (CGRP) α и β (37 аминокислот) экспрессируются в ряде нейронов ЦНС и периферической нервной системы (особенно в связи с кровеносными сосудами). Их функции - участие в ноцицепции, пищевом поведении, в регуляции тонуса ГМК сосудов (вазодилатация), бронхов (бронхоконстрикция).
Хюртля клетки
Иногда в составе стенки фолликулов или между фолликулами находят крупные клетки с зернистой оксифильной цитоплазмой, содержащие много митохондрий - онкоциты, или клетки Хюртля (Гюртля, также Асканази-Хюртля).
Межфолликулярные клетки
К паренхиме щитовидной железы, помимо образующих фолликулы клеток, относятся также островки клеток, расположенные между фолликулами. Островки образованы способными синтезировать йодсодержащие гормоны клетками (малодифференцированные тироциты, формирующие новые фолликулы), а также С-клетками.
СТРОМА
Строма состоит из вспомогательных структур (капсула, интерстиций, нервные и сосудистые элементы). Капсула сформирована из плотной волокнистой соединительной ткани. От капсулы отходят тяжи (стандартное наименование - септы, или трабекулы) плотной волокнистой соединительной ткани, содержащие кровеносные и лимфатические сосуды, нервы.
Интерстиций. Пространство органа заполняет поддерживающий элементы паренхимы каркас из рыхлой волокнистой соединительной ткани с кровеносными и лимфатическими сосудами, отдельными нервными волокнами и их окончаниями.
Кровоток железы интенсивен и сопоставим с кровоснабжением мозга, перфузией крови через почки и печень. Кровеносные капилляры фенестрированного типа контактируют с эндокринными клетками паренхимы.
Иннервация
Соматическая чувствительная. В железе найдены чувствительные нервные окончания, образованные ветвлениями периферических отростков чувствительных нейронов.
Двигательная вегетативная (симпатическая и парасимпатическая). Преобладают сопровождающие кровеносные сосуды и иннервирующие их ГМК варикозные ветвления постганглионарных симпатических нейронов. Эффекты вегетативной иннервации на эндокринную функцию незначительны.
ОКОЛОЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Четыре небольшие паращитовидные железы расположены на задней поверхности и под капсулой щитовидной железы. Эпителий нижних двух паращитовидных желёз развивается из энтодермы третьей пары глоточных карманов, верхних двух - из четвёртой пары. Функция желёз - синтез и секреция Са 2 +-регулирующего пептидного гормона паратиреокрина (паратиреоидного гормона, ПТГ). ПТГ вместе с кальцитонином и катакальцином, а также с витамином D регулирует обмен кальция и фосфатов.
Каждая из четырёх желёз имеет собственную тонкую капсулу, от которой отходят перегородки (септы), содержащие кровеносные сосуды. Паренхима, образованная тяжами и островками эпителиальных клеток, содержит два типа клеток - главные и оксифильные.
Главные клетки имеют базофильную цитоплазму (развита гранулярная эндоплазматическая сеть), комплекс Гольджи, мелкие митохондрии и секреторные гранулы диаметром 200-400 нм, содержащие
Оксифильные клетки равномерно распределены в паренхиме железы или образуют небольшие скопления, содержат крупные митохондрии, слабо выраженный комплекс Гольджи и умеренно развитую гранулярную эндоплазматическую сеть. Функция оксифильных клеток неизвестна, их число с возрастом увеличивается.
Жировые клетки всегда присутствуют в железе, с возрастом их количество увеличивается.
Паратиреоидный гормон, или паратиреокрин (паратирин, паратгормон, гормон паращитовидной железы, ПТГ, состоит из 84 аминокислотных остатков) поддерживает гомеостаз кальция и фосфатов. Регулятор экспрессии ПТГ - ионы Са 2+ , взаимодействующие с трансмембранными рецепторами главных клеток паращитовидных желёз. Са 2+ сыворотки регулирует секрецию ПТГ по механизму отрицательной обратной связи. Функции. ПТГ поддерживает гомеостаз Ca 2 +. Паратиреокрин увеличивает содержание Ca 2+ в плазме, усиливая его вымывание из костей, реабсорбцию в канальцах почки и всасывание в кишечнике.
НАДПОЧЕЧНИК
Надпочечники (см. рис. 9-24) - парные эндокринные органы, расположенные ретроперитонеально у верхних полюсов почки на уровне Th 12 и L 1 ; масса надпочечника - примерно 4 г. Фактически это две железы: кора (на долю коры приходится около 80% массы железы) и мозговая часть. Кора надпочечников синтезирует кортикостероиды (минералокортикоиды, глюкокортикоиды и андрогены), хромаффинная ткань мозговой части - катехоловые амины.
Развитие. На 6-й неделе внутриутробного развития крупные мезодермальные клетки целомического эпителия образуют скопления между основанием дорзальной брыжейки первичной кишки и развивающимися урогенитальными валиками. По направлению к этим скоплениям из ближайших симпатических ганглиев мигрируют клетки нервного гребня - будущие хромаффинные клетки мозгового вещества. В дальнейшем число хромаффинных клеток возрастает вплоть до завершения полового развития. Мезодермальные клетки формируют две зоны коры: наружную - дефинитивную и эмбриональную (фетальную), расположенную на границе с мозговым веществом. Незадолго до рождения начинается дегенерация фетальной коры, и к концу первого года жизни фетальная кора полностью исчезает. В течение первого года жизни в дефинитивной коре различимы клубочковая, пучковая и сетчатая зоны; полностью дифференцировка корковой части надпочечника завершается к третьему году жизни. Регенерация. Клетки коры и мозговой части железы способны поддерживать свою численность как путём их пролиферации, так и за счёт камбиального резерва.
Кора. Непосредственно под капсулой органа находятся эпителиальные камбиальные клетки, постоянно дифференцирующиеся в эндокринные клетки коры. АКТГ стимулирует пролиферацию камбиального резерва.
Мозговая часть. Часть мигрировавших сюда клеток нервного гребня сохраняется в виде камбиального резерва. Эти малодифференцированные клетки - источник развития новых хромаффинных клеток.
Кровоснабжение железы осуществляется из трёх источников: верхняя надпочечниковая артерия (ветвь нижней диафрагмальной артерии), средняя надпочечниковая артерия (отходит от аорты), нижняя надпочечниковая артерия (ветвь почечной артерии) (рис. 9-23). Верхняя и средняя надпочечниковые артерии дают начало капиллярам, пронизывающим корковое вещество и заканчивающимися мозговыми венозными синусами в мозговом веществе. Это означает, что гормоны, продуцируемые клетками коркового вещества, покидают кору, проходя через мозговое вещество, при этом глюкокортикоиды коры стимулируют секрецию адреналина из хромаффинных клеток. Это обстоятельство объясняет сочетанное вовлечение органа в развитие стрессовых ситуаций (адаптационный синдром, по Селье). Нижняя надпочечниковая артерия даёт начало мозговой артерии, которая кровоснабжает только мозговое вещество, минуя корковое, и заканчивается на мозговых венозных синусах. Медуллярные венозные синусы открываются в центральную вену.
КОРА НАДПОЧЕЧНИКА
Железа (рис. 9-24) окружена капсулой из плотной волокнистой соединительной ткани, от которой в толщу органа местами отходят соеди-
 Рис. 9-23. Кровоснабжение надпочечника.
Рис. 9-23. Кровоснабжение надпочечника.
нительнотканные перегородки. Строма железы состоит из поддерживающей эндокринные клетки рыхлой волокнистой соединительной ткани, содержащей огромное количество кровеносных капилляров с фенестированным эндотелием. Паренхима - совокупность эпителиальных тяжей, имеющих различную ориентацию на разном расстоянии от капсулы надпочечника. Это обстоятельство, а также характер гормонального стероидогенеза позволяет выделить в коре клубочковую, пучковую и сетчатую зоны.
Клубочковая зона. Тяжи эндокринных клеток подворачиваются под капсулу и на срезе имеют вид клубочков (15% толщины коры). Здесь синтезируются минералокортикоиды (преимущественно альдостерон). Стимулятор синтеза альдостерона - ангиотензин II и в незначительной степени - АКТГ. Клетки (рис. 9-25Б) имеют плотное округлое ядро с одним или двумя ядрышками, развитую гладкую эндоплазматическую сеть, некрупные митохондрии с пластинчатыми кристами, ри-
 Рис. 9-24. Надпочечник.
Непосредственно
под капсулой в составе корковой части находится клубочковая зона. Она
состоит из узких и более мелких по сравнению с другими зонами клеток.
Крупные многоугольные клетки образуют параллельные тяжи пучковой зоны.
Правильный ход тяжей нарушается в сетчатой зоне корковой части
надпочечника. Мозговая часть представлена переплетающимися тяжами
крупных хромаффинных клеток. К тяжам прилегают синусоидные кровеносные
капилляры с широким просветом.
Рис. 9-24. Надпочечник.
Непосредственно
под капсулой в составе корковой части находится клубочковая зона. Она
состоит из узких и более мелких по сравнению с другими зонами клеток.
Крупные многоугольные клетки образуют параллельные тяжи пучковой зоны.
Правильный ход тяжей нарушается в сетчатой зоне корковой части
надпочечника. Мозговая часть представлена переплетающимися тяжами
крупных хромаффинных клеток. К тяжам прилегают синусоидные кровеносные
капилляры с широким просветом.
босомы, хорошо развитый комплекс Гольджи и небольшое количество мелких липидных включений.
Пучковая зона занимает около 75% толщины коры. Тяжки эндокринных клеток и находящиеся между ними кровеносные капилляры расположены параллельно друг другу (в виде пучков). Здесь синтезируются глюкокортикоиды (преимущественно кортизол и кортизон), а также андрогены. Синтез глюкокортикоидов регулирует тропный гормон аденогипофиза - АКТГ. Клетки на гистологических препаратах выглядят как вакуолизированные (рис. 9-25А), поэтому их называют спонгиоциты. Вакуолизация клеток на гистологических препаратах отражает присутствие в цитоплазме спонгиоцитов значительного числа липидных капель (содержат преимущественно эфиры холестерина), вымываемых при подготовке препарата. Спонгиоциты содержат округлые митохондрии с кристами в виде трубочек и пузырьков, раз- ветвлённую гладкую эндоплазматическую сеть, элементы гранулярной эндоплазматической сети, лизосомы, многочисленные липидные включения и пигментные гранулы, содержащие липофусцин. Сетчатая зона. В наиболее глубоких частях коры (10% толщины коры) тяжи эндокринных клеток переплетаются, образуя подобие сети. В сетчатой зоне синтезируются глюкокортикоиды и стероидные гормоны типа андрогенов (дегидроэпиандростерон и андростендион). Тропный гормон - АКТГ. Гонадотропные гормоны гипофиза не влияют на секрецию гормонов в сетчатой зоне. В отличие от спонгиоцитов, клетки этой зоны содержат меньше липидных включений, но имеют крупные липофусциновые гранулы. Липофусциновые гранулы содержат лизосомальную кислую фосфатазу и рассматриваются как деградирующие лизосомы.
Стероидогенез гормонов коры надпочечника, а также стероидных гормонов половой сферы - сложный процесс (из железы выделено не менее 50 стероидов), по-разному происходящий в отдельных зонах коры. Стероидные гормоны, их промежуточные продукты, а также фармакологические аналоги гормонов синтезируются на базе холестерина. Процессы стероидогенеза обеспечивают ферменты, локализованные в митохондриях и гладкой эндоплазматической сети.
Глюкокортикоиды. Основной глюкокортикоид, секретируемый надпочечниками, - кортизол; на его долю приходится 80%. Остальные 20% - кортизон, кортикостерон, 11-дезоксикортизол и 11-дезоксикортикостерон. АКТГ - основной регулятор синтеза глюкокортикоидов. Для синтеза и секреции кортиколиберина, АКТГ и кортизола характерна выраженная суточная периодичность. При нормальном ритме сна увеличение секреции кортизола наступает после засыпания и достигает максимума при пробуждении. Функции глюкокортикоидов разнообразны: от регуляции метаболизма до модификации иммунологического и воспалительного ответов. Наиболее важный метаболический эффект глюкокортикоидов - преобразование жира и мышечных белков в гликоген.
 Рис. 9-25. Эндокринные клетки коры надпочечника.
А - клетка пучковой зоны, вырабатывающая глюкокортикоиды и андрогены. Клетку называют спонгиоцитом, т.к. она имеет пенистый
вид
из-за множества липидных капель в цитоплазме; содержит округлые
митохондрии с кристами в виде трубочек и пузырьков, разветвлённую
гладкую эндоплазматическую сеть. Б
- клетка клубочковой зоны,
вырабатывающая альдостерон. Присутствуют развитая гладкая
эндоплазматическая сеть, некрупные митохондрии с пластинчатыми кристами и
небольшое количество мелких липидных включений.
Рис. 9-25. Эндокринные клетки коры надпочечника.
А - клетка пучковой зоны, вырабатывающая глюкокортикоиды и андрогены. Клетку называют спонгиоцитом, т.к. она имеет пенистый
вид
из-за множества липидных капель в цитоплазме; содержит округлые
митохондрии с кристами в виде трубочек и пузырьков, разветвлённую
гладкую эндоплазматическую сеть. Б
- клетка клубочковой зоны,
вырабатывающая альдостерон. Присутствуют развитая гладкая
эндоплазматическая сеть, некрупные митохондрии с пластинчатыми кристами и
небольшое количество мелких липидных включений.
Минералокортикоиды. Альдостерон - основной минералокортикоид. Другие стероиды надпочечника - кортизол, 11-дезоксикортизол, 11-дезоксикортикостерон, кортикостерон - имеют и минералокортикоидную активность, хотя - сравнительно с альдостероном - их суммарный вклад мал. Ангиотензин II - компонент системы «ренин-ангиотензины» - главный регулятор синтеза и секреции альдостерона. Этот пептид стимулирует выброс альдостерона. Натриуретические факторы ингибируют синтез альдостерона. Функция минералокортикоидов - поддержание баланса электролитов жидкостей организма, осуществляется посредством влияния на реабсорбцию ионов в почечных канальцах.
Андрогены. В коре надпочечников синтезируются дегидроэпиандростерон и в меньшей степени андростендион.
МОЗГОВАЯ ЧАСТЬ НАДПОЧЕЧНИКА
Эндокринную функцию мозговой части надпочечника выполняют происходящие из нервного гребня хромаффинные клетки. При активации симпатической нервной системы надпочечники выбрасывают в кровь катехоловые амины (адреналин и норадреналин). Катехоламины имеют широкий спектр эффектов (воздействие на гликогенолиз, липолиз, глюконеогенез, существенное влияние на сердечно-сосудистую систему). Вазоконстрикция, параметры сокращения сердечной мышцы и другие эффекты катехоловых аминов реализуются через α- и β-адренергические рецепторы на поверхности клеток-мишеней (ГМК, секреторные клетки, кардиомиоциты). Серьёзные клинические проблемы возникают при опухолях эндокринных клеток и их предшественников (нейробластома, феохромоцитома). Строма. В нежном поддерживающем каркасе, состоящем из рыхлой волокнистой соединительной ткани, расположены многочисленные сосудистые полости - венозные синусы - вариант капилляров типа синусоидов. Их отличительная особенность - значительный диаметр просвета, достигающий десятков и сотен мкм.
Иннервация. Мозговая часть органа содержит множество преганглионарных нервных волокон симпатического отдела нервной системы, хромаффинные клетки расценивают как постганглионарное звено (модифицированные постганглионарные симпатические нейроны) двигательной вегетативной иннервации. Между хромаффинными клетками в мозговом веществе можно также видеть рассеянные небольшие группы ганглионарных клеток с неясной функцией.
Хромаффинные клетки
Хромаффинные клетки (рис. 9-29) содержат гранулы с электронноплотным содержимым, которое с бихроматом калия даёт хромаффинную реакцию. Хромаффинные клетки - основной клеточный элемент мозговой части надпочечников и параганглиев, расположенных
 Рис. 9-29. Хромаффинная клетка.
Характерны
многочисленные электронно-плотные гранулы с катехоламинами.
Значительный объём клетки занимает крупное ядро. Клетка содержит
митохондрии, выраженный комплекс Гольджи, элементы гранулярной
эндоплазматической сети.
Рис. 9-29. Хромаффинная клетка.
Характерны
многочисленные электронно-плотные гранулы с катехоламинами.
Значительный объём клетки занимает крупное ядро. Клетка содержит
митохондрии, выраженный комплекс Гольджи, элементы гранулярной
эндоплазматической сети.
по ходу крупных артериальных стволов (например, каротидное тело). Мелкие скопления и одиночные хромаффинные клетки находят также в сердце, почках, симпатических ганглиях.
Хромаффинные клетки содержат многочисленные митохондрии, выраженный комплекс Гольджи, элементы гранулярной эндоплазматической сети, многочисленные электронно-плотные гранулы, содержащие преимущественно норадреналин и/или адреналин (по этому признаку хромаффинные клетки подразделяют на две субпопуляции), а также АТФ, энкефалины и хромогранины. Адреналин-содержащие гранулы гомогенны. Норадреналин-содержащие гранулы характеризуются повышенной плотностью содержимого в центральной части и наличием светлого ободка по периферии под мембраной гранулы. Секреция гормонов из хромаффинных клеток происходит в результате стимулирующего влияния со стороны преганглионарных симпатических волокон и глюкокортикоидов. Секрет хромаффинных клеток содержит 10% норадреналина и 90% адреналина. Эти катехоламины имеют широкий спектр эффектов (воздействие на гликогенолиз, ли-
полиз, глюконеогенез, существенно влияние на сердечно-сосудистую систему). Вазоконстрикция, параметры сокращения сердечной мышцы и другие эффекты катехоловых аминов реализуются через α- и β-адренергические рецепторы на поверхности клеток-мишеней (ГМК, секреторные клетки, кардиомиоциты).
ЖЕЛЕЗЫ ВНУТРЕННЕЙ СЕКРЕЦИИ (ОБЩАЯ)
ü Понятие о железах внутренней секреции (ЖВС) сформулировано И. Мюллером (1830).
ü Немецкий физиолог Адольф Бертольд (1849) установил, что пересадка кастрированному петуху в брюшную полость семенников другого петуха приводит к восстановлению исходных свойств у кастрата.
ü В 1889 Броун –Секар сообщил об опытах, проведенных на самом себе –вытяжки из семенников животных оказали на старческий организм (ученому 72 года) «омолаживающее действие», но эффект омоложения длился недолго -через 2-3 месяца он пропадал.
ü В 1901 году Соболев Л.В., доказал секрецию панкреатической железой инсулина (1921 г. Бантинг и Ч. Бест).
Эндокринология –наука, изучающая развитие, строение, функции ЖВС и клеток –продуцентов гормонов, биосинтез, механизм действия и особенности гормонов, их секрецию в норме и при патологии, а так же болезни, возникшие в результате нарушения продукции гормонов.
ЖВС - это специализированные в процессе фило- и онтогенеза органы или группы клеток, синтезирующие и выделяющие во внутреннюю среду организма биологически активные вещества (БАВ) – гормоны. ЖВС не имеют выводных протоков. Их клетки оплетены обильной сетью кровеносных и лимфатических капилляров, и их БАВ выделяются непосредственно в кровь и лимфу.
ГОРМОНЫ
Гормоны - это группа высокоспециализированных БАВ, обеспечивающих регуляцию и интеграцию функций органов и всего организма в целом.
ФИЗИОЛОГИЧЕСКАЯ РОЛЬ ГОРМОНОВ В ОРГАНИЗМЕ:
1. Гомеостатическая функция.
2. Влияют на процессы роста , дифференцировки тканей(т. е. на физическое, умственное и половое созревание)
3. Обеспечивают адаптацию организма.
4. Регулируют репродуктивную функцию организма (оплодотворение, беременность, лактация).
5. Регулируют и интегрируют функции организма совместно с ЦНС.
Высшей формой гуморальной регуляции является гормональная . Термин "гормон " был впервые применен в 1902 г. Старлингом и Бейлиссом в отношении открытого ими вещества, продуцирующегося в двенадцатиперстной кишке, -секретина . Термин"гормон " в переводе с греческого означает "побуждающий к действию ", хотя не все гормоны обладают стимулирующим эффектом.
Классификация вариантов действия гормонов (Балаболкин М.М., 1989):
1. Гормональное (или собственно эндокринное) - гормон выделяется из клетки –продуцента, попадает в кровь и с током крови подходит к органу –мишени, действуя на расстоянии от места продукции гормона.
2. Паракринное - из места синтеза гормон попадает во внеклеточное пространство, из него –воздействует на клетки –мишени, расположенные в округе(простагландины).
3. Аутокринное - клетки продуцируют гормон, который сам и воздействует на эту же клетку –продуцент, то есть клетка –мишень = клетка –продуцент.
ОТЛИЧИТЕЛЬНЫЕ ОСОБЕННОСТИ ГОРМОНОВ:
1.Имеют высокую биологическую активность (мг, нг).
2.Секреция гормона - путем экзоцитоза.
3.Гормоны поступают непосредственно в кровь, лимфу или окружающую секреторную клетку интерстициальную жидкость.
4.Гормон обладает дистантностью действия.
5.Гормон обладает высокой специфичностью действия, то есть вызывает строго специфичные ответы определенных органов или тканей-мишеней. В то же время клетки других тканей не реагируют на наличие гормона.
6.Гормон не служит источником энергии для клетки.
Гормоны синтезируются и выделяются тканями, не относящимся к железам внутренней секреции:
- жировой тканью , которая выделяет женские половые гормоны;
- миокардом , выделяющим натрийуретический гормон;
- слюнными железами - эпидермальный фактор роста;
- печенью, мышцами - инсулиноподобные соматомедины.
ВИДЫ ВОЗДЕЙСТВИЙ ГОРМОНОВ НА КЛЕТКИ-МИШЕНИ:
1. Прямое воздействие: гормон непосредственно сам вызывает изменения в клетках или тканях, органах.
2. Пермиссивное воздействие: через облегчения воздействия другого гормона на данную ткань. Например, глюкокортикоиды, сами не влияя на тонус мускулатуры сосудов, создают условия, для адреналина, который увеличивает АД.
3. Сенсибилизирующее воздействие: повышение чувствительности ткани к действию гормонов.
4. Синергическое воздействие: один гормон усиливает эффект другого гормона. Например, однонаправленное действие адреналина и глюкагона. Оба гормона активируют распад гликогена в печени до глюкозы и вызывают увеличение сахара в крови.
5. Антогонистическое воздействие. Так, инсулин и адреналин оказывают на уровень глюкозы крови противоположенное влияние: инсулин вызывает гипогликемию, а глюкагон - гипергликемию.
КЛАССИФИКАЦИЯ ГОРМОНОВ
1.По месту действия:
эффекторные гормоны: действуют непосредственно на органы-мишени;
тропные гормоны: действуют на другие эндокринные железы;
гипоталамические факторы (рилизинг-факторы) : действуют на гипофиз
Ø высвобождающие (либерины)
Ø ингибирующие (статины).
2. По биологическим функциям:
Гомеостаз жидкости и электролитов: АДГ, альдостерон, ангиотензин, натрийуретический гормон;
Регуляция Са: паратиреоидный гормон, кальцитонин, витамин Д.
Биологически активное вещество (БАВ), физиологически активное вещество (ФАВ) - вещество, которое в малых количествах (мкг, нг) оказывает выраженный физиологический эффект на различные функции организма.
Гормон — физиологически активное вещество, вырабатываемое или специализированными эндокринными клетками, выделяемое во внутреннюю среду организма (кровь, лимфа) и оказывающее дистантное действие на клетки-мишени.
Гормон - это сигнальная молекула, секретируемая эндокринными клетками, которая посредством взаимодействия со специфическими рецепторами клеток-мишеней регулирует их функции. Поскольку гормоны являются носителями информации, то они, как и другие сигнальные молекулы, обладают высокой биологической активностью и вызывают ответные реакции клеток-мишеней в очень малых концентрациях (10 -6 — 10 -12 М/л).
Клетки-мишени (ткани-мишени, органы-мишени) — клетки, ткани или органы, в которых имеются специфичные для данного гормона рецепторы. Некоторые гормоны имеют единственную ткань-мишень, тогда как другие оказывают влияние повсеместно в организме.
Таблица. Классификация физиологически активных веществ
Свойства гормонов
Гормоны имеют ряд общих свойств. Обычно они образуются специализированными эндокринными клетками. Гормоны обладают избирательностью действия, которая достигается благодаря связыванию со специфическими рецепторами, находящимися на поверхности клеток (мембранные рецепторы) или внутри них (внутриклеточные рецепторы), и запуску каскада процессов внутриклеточной передачи гормонального сигнала.
Последовательность событий передачи гормонального сигнала может быть представлена в виде упрощенной схемы «гормон (сигнал, лиганд) -> рецептор -> второй (вторичный) посредник -> эффекторные структуры клетки -> физиологический ответ клетки». У большинства гормонов отсутствует видовая специфичность (за исключением ), что позволяет изучать их эффекты на животных, а также использовать гормоны, полученные от животных, для лечения больных людей.
Различают три варианта межклеточного взаимодействия с помощью гормонов:
- эндокринный (дистантный), когда они доставляются к клеткам-мишеням от места продукции кровью;
- паракринный — гормоны диффундируют к клетке-мишени от рядом расположенной эндокринной клетки;
- аутокринный — гормоны воздействуют на клетку-продуцент, которая одновременно является для него клеткой-мишенью.
По химической структуре гормоны делят на три группы:
- пептиды (число аминокислот до 100, например тиротропина рилизинг-гормон, АКТГ) и белки (инсулин, гормон роста, и др.);
- производные аминокислот: тирозина (тироксин, адреналин), триптофана — мелатонин;
- стероиды, производные холестерола (женские и мужские половые гормоны, альдостерон, кортизол, кальцитриол) и ретиноевая кислота.
По выполняемой функции гормоны делят на три группы:
- эффекторные гормоны , действующие непосредственно на клетки-мишени;
- тронные гормоны гипофиза , контролирующие функцию периферических эндокринных желез;
- гормоны гипоталамуса , регулирующие секрецию гормонов гипофизом.
Таблица. Типы действия гормонов
|
Тип действия |
Характеристика |
|
Гормональное (гемокринное) |
Действие гормона на значительном удалении от места образования |
|
Изокринное (местное) |
Гормон, синтезируемый в одной клетке, оказывает действие на клетку, расположенную в тесном контакте с первой. Его высвобождение осуществляется в межтканевую жидкость и кровь |
|
Нейрокринное (нейроэндокринное) |
Действие, когда гормон, высвобождаясь из нервных окончаний, выполняет функцию нейромедиатора или нейромодулятора |
|
Паракринное |
Разновидность изокринного действия, но при этом гормон, образующийся в одной клетке, поступает в межклеточную жидкость и влияет на ряд клеток, расположенных в непосредственной близости |
|
Юкстакринное |
Разновидность паракринного действия, когда гормон не попадает в межклеточную жидкость, а сигнал передастся через плазматическую мембрану рядом расположенной клетки |
|
Аутокринное |
Высвобождающийся из клетки гормон оказывает влияние на ту же клетку, изменяя ее функциональную активность |
|
Соликринное |
Высвобождающийся из клетки гормон поступает в просвет протока и достигает, таким образом, другой клетки, оказывая на нес специфическое воздействие (характерно для желудочно- кишечных гормонов) |
Гормоны циркулируют в крови в свободном (активная форма) и связанном (неактивная форма) состоянии с белками плазмы или форменных элементов. Биологической активностью обладают гормоны в свободном состоянии. Содержание их в крови зависит от скорости секреции, степени связывания, захвата и скорости метаболизма в тканях (связывания со специфическими рецепторами, разрушения или инактивации в клетках-мишенях или гепатоцитах), удаления с мочой или желчью.
Таблица. Физиологически активные вещества, открытые в последнее время

Ряд гормонов может подвергаться в клетках-мишенях химическим превращениям в более активные формы. Так, гормон «тироксин», подвергаясь дейодированию, превращается в более активную форму — трийодтиронин. Мужской половой гормон тестостерон в клетках-мишенях может не только превращаться в более активную форму — дегидротестостерон, но и в женские половые гормоны группы эстрогенов.
Действие гормона на клетку-мишень обусловлено связыванием, стимуляцией специфического к нему рецептора, после чего происходит передача гормонального сигнала на внутриклеточный каскад превращений. Передача сигнала сопровождается его многократным усилением, и действие на клетку небольшого числа молекул гормона может сопровождаться мощной ответной реакцией клеток-мишеней. Активация гормоном рецептора сопровождается также включением внутриклеточных механизмов, прекращающих ответ клетки на действие гормона. Это могут быть механизмы, понижающие чувствительность (десенситизация/адаптация) рецептора к гормону; механизмы, дефосфорилирующие внутриклеточные ферментные системы и др.
Рецепторы к гормонам, как и к другим сигнальным молекулам, локализованы на клеточной мембране или внутри клетки. С рецепторами клеточной мембраны (1-TMS, 7-TMS и лигандзависимые ионные каналы) взаимодействуют гормоны гидрофильной (лииофобной) природы, для которых клеточная мембрана не проницаема. Ими являются катехоламины, мелатонин, серотонин, гормоны белково-пептидной природы.
Гормоны гидрофобной (липофильной) природы диффундируют через плазматическую мембрану и связываются с внутриклеточными рецепторами. Эти рецепторы делятся на цитозольные (рецепторы стероидных гормонов — глюко- и минералокортикоидов, андрогенов и прогестинов) и ядерные (рецепторы тиреоидных йодсодержащих гормонов, кальцитриола, эстрогенов, ретиноевой кислоты). Цитозольные рецепторы и рецепторы эстрогенов связаны с белками теплового шока (БТШ), что предотвращает их проникновение в ядро. Взаимодействие гормона с рецептором приводит к отделению БТШ, образованию гормон-рецепторного комплекса и активации рецептора. Комплекс гормон-рецептор перемещается в ядро, где он взаимодействует со строго определенными гормон-чувствительными (узнающими) участками ДНК. Это сопровождается изменением активности (экспрессией) определенных генов, контролирующих синтез белков в клетке и другие процессы.
По использованию тех или иных внутриклеточных путей передачи гормонального сигнала наиболее распространенные гормоны можно разделить на ряд групп (табл. 8.1).
Таблица 8.1. Внутриклеточные механизмы и пути действия гормонов

Гормоны контролируют разнообразные реакции клеток-мишеней и через них — физиологические процессы организма. Физиологические эффекты гормонов зависят от их содержания в крови, количества и чувствительности рецепторов, состояния пострецепторных структур в клетках-мишенях. Под действием гормонов может происходить активация или торможение энергетического и пластического метаболизма клеток, синтеза различных, в том числе белковых веществ (метаболическое действие гормонов); изменение скорости деления клетки, ее дифференцировки (морфогенетическое действие), инициирование запрограммированной гибели клетки (апоптоз); запуск и регуляция сокращения и расслабления гладких миоцитов, секреции, абсорбции (кинетическое действие); изменение состояния ионных каналов, ускорение или торможение генерации электрических потенциалов в водителях ритма (корригирующее действие), облегчение или угнетение влияния других гормонов (реактогенное действие) и т.д.
Таблица. Распределение гормона в крови

Скорость возникновения в организме и продолжительность ответных реакций на действие гормонов зависит от типа стимулируемых рецепторов и скорости метаболизма самих гормонов. Изменения физиологических процессов могут наблюдаться через несколько десятков секунд и длиться кратковременно при стимуляции рецепторов плазматической мембраны (например, сужение сосудов и повышение артериального давления крови под действием адреналина) или наблюдаться через несколько десятков минут и длиться часами при стимуляции ядерных рецепторов (например, усиление обмена в клетках и увеличение потребления кислорода организмом при стимуляции тиреоидных рецепторов трийодтиронином).
Таблица. Время действия физиологически активных веществ
Поскольку одна и та же клетка может содержать рецепторы к разным гормонам, то она способна быть одновременно клеткой-мишенью для нескольких гормонов и других сигнальных молекул. Действие одного гормона на клетку нередко сочетается с влиянием других гормонов, медиаторов, цитокинов. При этом в клетках-мишенях может происходить запуск ряда путей передачи сигналов, в результате взаимодействия которых может наблюдаться усиление или торможение ответной реакции клетки. Например, на гладкий миоцит стенки сосудов могут одновременно действовать норадреналин и , суммируя их сосудосуживающее влияние. Сосудосуживающее действие вазопрессина может быть устранено или ослаблено одновременным действием на гладкие миоциты сосудистой стенки брадикинина или оксида азота.
Регуляция образования и секреции гормонов
Регуляция образования и секреции гормонов является одной из важнейших функций и нервной систем организма. Среди механизмов регуляции образования и секреции гормонов выделяют влияние ЦНС, «тройных» гормонов, влияние по каналам отрицательной обратной связи концентрации гормонов в крови, влияние конечных эффектов гормонов на их секрецию, влияние суточных и других ритмов.
Нервная регуляция осуществляется в различных эндокринных железах и клетках. Это регуляция образования и секреции гормонов нейросекреторными клетками переднего гипоталамуса в ответ на поступление к нему нервных импульсов с различных областей ЦНС. Эти клетки обладают уникальной способностью возбуждаться и трансформировать возбуждение в образование и секрецию гормонов, стимулирующих (рилизинг-гормоны, либерины) или тормозящих (статины) секрецию гормонов гипофизом. Например, при увеличении притока нервных импульсов к гипоталамусу в условиях психоэмоционального возбуждения, голода, болевого воздействия, действии тепла или холода, при инфекции и в других чрезвычайных условиях, нейросекреторные клетки гипоталамуса высвобождают в портальные сосуды гипофиза кортикотропина рилизинг-гормон, который усиливает секрецию адренокортикотропного гормона (АКТГ) гипофизом.
Непосредственное влияние на образование и секрецию гормонов оказывает АНС. При повышении тонуса СНС увеличивается секреция тройных гормонов гипофизом, секреция катехоламинов мозговым веществом надпочечников, тиреоидных гормонов щитовидной железой, снижается секреция инсулина. При повышении тонуса ПСНС увеличивается секреция инсулина, гастрина и тормозится секреция тиреоидных гормонов.
Регуляции тронными гормонами гипофиза используется для контроля образования и секреции гормонов периферическими эндокринными железами (щитовидной, корой надпочечников, половыми железами). Секреция тропных гормонов находится под контролем гипоталамуса. Тропные гормоны получили свое название из-за их способности связываться (обладать сродством) с рецепторами клеток-мишеней, формирующих отдельные периферические эндокринные железы. Троп- ный гормон к тироцитам щитовидной железы называют тиро- тропином или тиреотропным гормоном (ТТГ), к эндокринным клеткам коры надпочечников — адренокортикотропным гормоном (АКГТ). Тропные гормоны к эндокринным клеткам половых желез получили название: лютропин или лютеинизирующий гормон (ЛГ) — к клеткам Лейдига, желтому телу; фоллитропин или фолликулостимулирующий гормон (ФСГ) — к клеткам фолликулов и клеткам Сертоли.
Тропные гормоны при повышении их уровня в крови многократно стимулируют секрецию гормонов периферическими эндокринными железами. Они могут оказывать на них также другие эффекты. Так, например, ТТГ усиливает в щитовидной железе кровоток, активирует метаболические процессы в тироцитах, захват ими йода из крови, ускоряет процессы синтеза и секреции тиреоидных гормонов. При избыточном количестве ТТГ наблюдается гипертрофия щитовидной железы.
Регуляция обратными связями используется для контроля секреции гормонов гипоталамуса и гипофиза. Ее суть заключается в том, что нейросекреторные клетки гипоталамуса имеют рецепторы и являются клетками-мишенями гормонов периферической эндокринной железы и тройного гормона гипофиза, контролирующего секрецию гормонов этой периферической железой. Таким образом, если под влиянием гипоталамического тиреотропин-рилизинг-гормона (ТРГ) увеличится секреция ТТГ, то последний свяжется не только с рецепторами тирсоцитов, но и с рецепторами нейросекреторных клеток гипоталамуса. В щитовидной железе ТТГ стимулирует образование тиреоидных гормонов, а в гипоталамусе — тормозит дальнейшую секрецию ТРГ. Связь между уровнем ТТГ в крови и процессами образования и секреции ТРГ в гипоталамусе получила название короткой петли обратной связи.
На секрецию ТРГ в гипоталамусе оказывает влияние и уровень гормонов щитовидной железы. Если их концентрация в крови повышается, то они связываются с рецепторами тиреоидных гормонов нейросекреторных клеток гипоталамуса и тормозят синтез и секрецию ТРГ. Связь между уровнем тиреоидных гормонов в крови и процессами образования и секреции ТРГ в гипоталамусе получила название длинной петли обратной связи. Имеются экспериментальные данные о том, что гормоны гипоталамуса не только регулируют синтез и выделение гормонов гипофиза, но и тормозят собственное выделение, что определяют понятием сверхкороткой петли обратной связи.
Совокупность железистых клеток гипофиза, гипоталамуса и периферических эндокринных желез и механизмов их взаимного влияния друг на друга назвали системами или осями гипофиз — гипоталамус — эндокринная железа. Выделяют системы (оси) гипофиз — гипоталамус — щитовидная железа; гипофиз — гипоталамус — кора надпочечников; гипофиз — гипоталамус — половые железы.
Влияние конечных эффектов гормонов на их секрецию имеет место в островковом аппарате поджелудочной железы, С-клетках щитовидной железы, паращитовидных железах, гипоталамусе и др. Это демонстрируется следующими примерами. При повышении в крови уровня глюкозы стимулируется секреция инсулина, а при понижении — глюкагона. Эти гормоны по паракринному механизму тормозят секрецию друг друга. При повышении в крови уровня ионов Са 2+ стимулируется секреция кальцитонина, а при понижении — паратирина. Прямое влияние концентрации веществ на секрецию гормонов, контролирующих их уровень, является быстрым и эффективным способом поддержания концентрации этих веществ в крови.
Среди рассматриваемых механизмов регуляции секреции гормонов их конечными эффектами можно отметить регуляцию секреции антидиуретического гормона (АДГ) клетками заднего гипоталамуса. Секреция этого гормона стимулируется при повышении осмотического давления крови, например при потере жидкости. Снижение диуреза и задержка жидкости в организме под действием АДГ ведут к снижению осмотического давления и торможению секреции АДГ. Похожий механизм используется для регуляции секреции натрийуретического пептида клетками предсердий.
Влияние суточных и других ритмов на секрецию гормонов имеет место в гипоталамусе, надпочечниках, половых, шишковидной железах. Примером влияния суточного ритма является суточная зависимость секреции АКТГ и кортикостероидных гормонов. Самый низкий их уровень в крови наблюдается в полночь, а самый высокий — утром после пробуждения. Наиболее высокий уровень мелатонина регистрируется ночью. Хорошо известно влияние лунного цикла на секрецию половых гормонов у женщин.
Определение гормонов
Секреция гормонов - поступление гормонов во внутреннюю среду организма. Полипептидные гормоны накапливаются в гранулах и секретируются путем экзоцитоза. Стероидные гормоны не накапливаются в клетке и секретируются сразу после синтеза путем диффузии через клеточную мембрану. Секреция гормонов в большинстве случаев имеет циклический, пульсирующий характер. Периодичность секреции — от 5-10 мин до 24 ч и более (распространенный ритм — около 1 ч).
Связанная форма гормона — образование обратимых, соединенных нековалентными связями комплексов гормонов с белками плазмы и форменными элементами. Степень связывания различных гормонов сильно варьирует и определяется их растворимостью в плазме крови и наличием транспортного белка. Например, 90 % кортизола, 98 % тестостерона и эстрадиола, 96 % трийодтиронина и 99 % тироксина связываются с транспортными белками. Связанная форма гормона не может взаимодействовать с рецепторами и формирует резерв, который может быть быстро мобилизован для пополнения пула свободного гормона.
Свободная форма гормона — физиологически активное вещество в плазме крови в несвязанном с белком состоянии, способное взаимодействовать с рецепторами. Связанная форма гормона находится в динамическом равновесии с пулом свободного гормона, который в свою очередь находится в равновесии с гормоном, связанным с рецепторами в клетках-мишенях. Большинство полипептидных гормонов, за исключением соматотропина и окситоцина, циркулирует в низких концентрациях в крови в свободном состоянии, не связываясь с белками.
Метаболические превращения гормона - его химическая модификация в тканях-мишенях или других образованиях, обусловливающая снижение/повышение гормональной активности. Важнейшим местом обмена гормонов (их активации или инактивации) является печень.
Скорость метаболизма гормона - интенсивность его химического превращения, которая определяет длительность циркуляции в крови. Период полураспада катехоламинов и полипептидных гормонов составляет несколько минут, а тиреоидных и стероидных гормонов — от 30 мин до нескольких суток.
Гормональный рецептор — высокоспециализированная клеточная структура, входящая в состав плазматических мембран, цитоплазмы или ядерного аппарата клетки и образующая специфичное комплексное соединение с гормоном.
Органоспецифичность действия гормона - ответные реакции органов и тканей на физиологически активные вещества; они строго специфичны и не могут быть вызваны другими соединениями.
Обратная связь — влияние уровня циркулирующего гормона на его синтез в эндокринных клетках. Длинная цепь обратной связи — взаимодействие периферической эндокринной железы с гипофизарными, гипоталамическими центрами и с супрагипоталамическими областями ЦНС. Короткая цепь обратной связи — изменение секреции гипофизарного тронного гормона, модифицирует секрецию и высвобождение статинов и либеринов гипоталамуса. Ультракороткая цепь обратной связи — взаимодействие в пределах эндокринной железы, при котором выделение гормона влияет на процессы секреции и высвобождения его самого и других гормонов из данной железы.
Отрицательная обратная связь - повышение уровня гормона, приводящее к торможению его секреции.
Положительная обратная связь — повышение уровня гормона, обусловливающее стимуляцию и возникновение пика его секреции.
Анаболические гормоны -
физиологически активные вещества, способствующие образованию и обновлению структурных частей организма и накоплению в нем энергии. К таким веществам относятся гонадотропные гормоны гипофиза (фоллитропин, лютропин), половые стероидные гормоны (андрогены и эстрогены), гормон роста (соматотропин), хориони- ческий гонадотропин плаценты, инсулин.
Инсулин — белковое вещество, вырабатываемое в β-клетках островков Лангерганса, состоящее из двух полипептидных цепей (А-цепь — 21 аминокислота, В-цепь — 30), снижающее уровень глюкозы крови. Первый белок, у которого была полностью определена первичная структура Ф. Сенгером в 1945-1954 гг.
Катаболические гормоны — физиологически активные вещества, способствующие распаду различных веществ и структур организма и высвобождению из него энергии. К таким веществам относятся кортикотропин, глюкокортикоиды (корти- зол), глюкагон, высокие концентрации тироксина и адреналина.
Тироксин (тетрайодтиронин) - йодсодержащее производное аминокислоты тирозина, вырабатываемое в фолликулах щитовидной железы, повышающее интенсивность основного обмена, теплопродукцию, оказывающее влияние на рост и дифференцировку тканей.
Глюкагон - полипептид, вырабатываемый в а-клетках островков Лангерганса, состоящий из 29 аминокислотных остатков, стимулирующий распад гликогена и повышающий уровень глюкозы крови.
Кортикостероидные гормоны - соединения, образующиеся в корковом веществе надпочечников. В зависимости от числа атомов углерода в молекуле делят на С 18 -стероиды — женские половые гормоны — эстрогены, С 19 -стероиды — мужские половые гормоны — андрогены, С 21 -стероиды — собственно кортикостероидные гормоны, обладающие специфическим физиологическим действием.
Катехоламины — производные пирокатехина, активно участвующие в физиологических процессах в организме животных и человека. К катехоламинам относятся адреналин, норадреналин и дофамин.
Симпатоадреналовая система — хромаффинные клетки мозгового вещества надпочечников и иннервирующие их преганглионарные волокна симпатической нервной системы, в которых синтезируются катехоламины. Хромаффинные клетки также обнаружены в аорте, каротидном синусе, внутри и около симпатических ганглиев.
Биогенные амины — группа азотсодержащих органических соединений, образующихся в организме путем декарбоксилирования аминокислот, т.е. отщепления от них карбоксильной группы — СООН. Многие из биогенных аминов (гистамин, серотонин, норадреналин, адреналин, дофамин, тирамин и др.) оказывают выраженный физиологический эффект.
Эйкозаноиды - физиологически активные вещества, производные преимущественно арахидоновой кислоты, оказывающие разнообразные физиологические эффекты и подразделяющиеся на группы: простагландины, простациклины, тром- боксаны, левугландины, лейкотриены и др.
Регуляторные пептиды — высокомолекулярные соединения, представляющие собой цепочку аминокислотных остатков, соединенных пептидной связью. Регуляторные пептиды, насчитывающие до 10 аминокислотных остатков, называют олигопептидами, от 10 до 50 — полипептидами, свыше 50 — белками.
Антигормон — защитное вещество, вырабатываемое организмом при длительном введении белковых гормональных препаратов. Образование антигормона является иммунологической реакцией на введение извне чужеродного белка. По отношению к собственным гормонам организм не образует антигормоны. Однако могут быть синтезированы вещества, близкие по строению к гормонам, которые при введении в организм действуют как антиметаболиты гормонов.
Антиметаболиты гормонов
— физиологически активные соединения, близкие по строению к гормонам и вступающие с ними в конкурентные, антагонистические отношения. Антиметаболиты гормонов способны занимать их место в физиологических процессах, совершающихся в организме, или блокировать гормональные рецепторы.
Тканевой гормон (аутокоид, гормон местного действия) — физиологически активное вещество, вырабатываемое неспециализированными клетками и оказывающее преимущественно местный эффект.
Нейрогормон — физиологически активное вещество, вырабатываемое нервными клетками.
Эффекторный гормон - физиологически активное вещество, оказывающее непосредственный эффект на клетки и органы-мишени.
Тронный гормон — физиологически активное вещество, действующее на другие эндокринные железы и регулирующее их функции.






