Legge del gradiente
Gli stimoli sono caratterizzati non solo dalla forza e dalla durata dell'azione, ma anche dalla velocità con cui la forza d'influenza sull'oggetto aumenta nel tempo, cioè pendenza.
È stata determinata la relazione tra la rapidità dell'aumento dell'intensità della stimolazione e l'entità dell'eccitazione Vlegge del gradiente:la reazione di un sistema vivente dipende dal gradiente di stimolazione: maggiore è la ripidità dell'aumento dello stimolo nel tempo, maggiore, entro certi limiti, l'entità della risposta funzionale. IN vista generale La base fisiologica della legge del gradiente può essere presentata come segue.
Una diminuzione della pendenza dell'aumento della forza dello stimolo porta ad un aumento della soglia di eccitazione, a seguito della quale la risposta del biosistema ad una certa pendenza minima scompare del tutto. Questo fenomeno si chiama alloggio.
17. La soglia di irritazione è la forza minima dello stimolo alla quale si verifica l'eccitazione.
b) La reobase è la forza minima dello stimolo che provoca l'eccitazione quando agisce per un tempo indefinitamente lungo. In pratica soglia e reobase hanno lo stesso significato. Più bassa è la soglia di irritazione o più bassa è la reobase, maggiore è l'eccitabilità del tessuto.
V) Tempo utile - tempo minimo azione di uno stimolo con una forza di una reobase durante la quale si verifica l'eccitazione.
d) La cronassia è il tempo minimo di azione di uno stimolo con una forza di due reobasi richiesto perché si verifichi l'eccitazione.
18. FISIOLOGIA DEL TESSUTO MUSCOLARE
Muovere il corpo nello spazio, mantenere una certa postura, il lavoro del cuore e dei vasi sanguigni e tratto digerente nell'uomo e nei vertebrati sono eseguiti da muscoli di due tipi principali: striati (scheletrici, cardiaci) e lisci, che differiscono tra loro per organizzazione cellulare e tissutale, innervazione e, in una certa misura, meccanismi di funzionamento. Allo stesso tempo, ci sono molte somiglianze nei meccanismi molecolari della contrazione muscolare tra questi tipi di muscoli.
Funzioni e proprietà dei muscoli scheletrici
I muscoli scheletrici sono parte integrante del sistema muscolo-scheletrico umano. In questo caso, i muscoli svolgono le seguenti funzioni:
1) fornire una certa postura del corpo umano;
2) muovere il corpo nello spazio;
3) spostare le singole parti del corpo l'una rispetto all'altra;
4) sono una fonte di calore, svolgendo una funzione termoregolatrice.
Il muscolo scheletrico ha le seguenti proprietà essenziali:
1) eccitabilità - la capacità di rispondere a uno stimolo modificando la conduttività ionica e il potenziale di membrana. IN condizioni naturali questo stimolo è il trasmettitore acetilcolina, che viene rilasciato nelle terminazioni presinaptiche degli assoni dei motoneuroni. In condizioni di laboratorio viene spesso utilizzata la stimolazione muscolare elettrica. Con la stimolazione elettrica del muscolo, inizialmente vengono eccitate le fibre nervose che rilasciano acetilcolina, cioè in questo caso si osserva un'irritazione indiretta del muscolo. Ciò è dovuto al fatto che l'eccitabilità delle fibre nervose è superiore a quella delle fibre muscolari. Per l'irritazione muscolare diretta, è necessario utilizzare miorilassanti - sostanze che bloccano la trasmissione degli impulsi nervosi attraverso la sinapsi neuromuscolare;
2) conduttività - la capacità di condurre un potenziale d'azione lungo e in profondità nella fibra muscolare lungo il sistema T;
3) contrattilità: la capacità di accorciare o sviluppare tensione quando eccitato;
4) elasticità: la capacità di sviluppare tensione quando allungato.
4. Organizzazione strutturale della fibra muscolare
Una fibra muscolare è una struttura multinucleare circondata da una membrana e contenente un apparato contrattile specializzato: le miofibrille. Oltretutto, componenti essenziali le fibre muscolari sono i mitocondri, un sistema di tubuli longitudinali - il reticolo sarcoplasmatico (reticolo) e un sistema di tubuli trasversali - il sistema T. Unità funzionale dell'apparato contrattile cellula muscolareè un sarcomero, la miofibrilla è costituita da sarcomeri.
Meccanismi di contrazione delle fibre muscolari
Nelle fibre muscolari a riposo, in assenza di impulsi dei motoneuroni, i ponti trasversali di miosina non sono attaccati ai miofilamenti di actina. La tropomiosina è posizionata in modo tale da bloccare le aree di actina che possono interagire con i ponti trasversali della miosina. La troponina inibisce l'attività della miosina-ATPasi e quindi l'ATP non viene scomposto. Le fibre muscolari sono in uno stato rilassato.
Quando un muscolo si contrae, la lunghezza dei dischi A non cambia, i dischi J si accorciano e la zona H dei dischi A può scomparire. Questi dati sono stati la base per creare una teoria che spiega la contrazione muscolare mediante scorrimento meccanismo (teoria dello scorrimento) miofilamenti sottili di actina lungo quelli spessi di miosina. Di conseguenza, i miofilamenti di miosina vengono retratti tra quelli circostanti di actina. Ciò porta all'accorciamento di ciascun sarcomero e quindi dell'intera fibra muscolare.
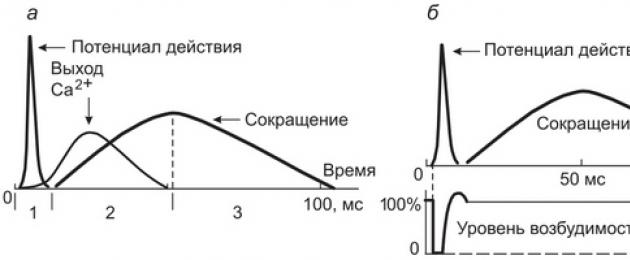
Meccanismo molecolare di contrazione fibra muscolare è che il potenziale d'azione derivante nell'area della placca terminale si propaga attraverso il sistema di tubuli trasversali in profondità nella fibra, provocando la depolarizzazione delle membrane dei serbatoi del reticolo sarcoplasmatico e il rilascio da esse ioni calcio. Gli ioni calcio liberi nello spazio interfibrillare innescano il processo di contrazione. L'insieme dei processi che causano la propagazione del potenziale d'azione in profondità nella fibra muscolare, il rilascio di ioni calcio dal reticolo sarcoplasmatico, l'interazione delle proteine contrattili e l'accorciamento della fibra muscolare è chiamato "accoppiamento elettromeccanico". La sequenza temporale tra la comparsa del potenziale d'azione delle fibre muscolari, l'ingresso degli ioni calcio nelle miofibrille e lo sviluppo della contrazione delle fibre è mostrata nella Figura 4.4.
Quando la concentrazione di ioni Ca 2+ nello spazio intermiofibrillare è inferiore a 10″, la tropomiosina si trova in modo tale da bloccare l'attaccamento dei ponti trasversali della miosina ai filamenti di actina. I ponti trasversali della miosina non interagiscono con i filamenti di actina. Non c'è movimento dei filamenti di actina e di miosina l'uno rispetto all'altro. Pertanto, la fibra muscolare è in uno stato rilassato. Quando la fibra è eccitata, il Ca 2+ esce dalle cisterne del reticolo sarcoplasmatico e, di conseguenza, aumenta la sua concentrazione in prossimità delle miofibrille. Sotto l'influenza dell'attivazione degli ioni Ca 2+, la molecola di troponina cambia forma in modo tale da spingere la tropomiosina nel solco tra i due filamenti di actina, liberando così siti per l'attacco dei ponti trasversali della miosina all'actina. Di conseguenza, i ponti trasversali sono attaccati ai filamenti di actina. Poiché le teste di miosina effettuano movimenti di “rematura” verso il centro del sarcomero, i miofilamenti di actina vengono “retratti” negli spazi tra i filamenti spessi di miosina e il muscolo si accorcia.
Il sistema muscolare ha una serie di proprietà fisiche e fisiologiche. Al principale Proprietà fisiche relazionare:
Birifrangenza(anisotropia). Formata dai dischi A, sta nel fatto che nella luce ordinaria le parti anisotrope appaiono scure, e nella luce polarizzata appaiono chiare se la luce viene trasmessa in direzione longitudinale, e scure se passa in direzione trasversale. L'alternanza di dischi anisotropi e isotropi crea striature trasversali nei muscoli.
Estensibilità. Associato alla presenza nei muscoli della componente elastica della membrana, striscia, reticolo sarcoplasmatico, ...).
Elasticità. Questa proprietà è associata all'estensibilità e sta nel fatto che dopo l'allungamento il muscolo ritorna nella sua posizione originale.
Elasticità. Questa proprietà del muscolo è associata alla sua compressione. Dopo la compressione, il muscolo è in grado di ritornare al suo stato originale.
Plastica. Consiste nel fatto che il muscolo è in grado di mantenere per qualche tempo la forma artificiale che gli è stata data. Le proprietà plastiche dei muscoli scheletrici sono espresse molto debolmente, sono più caratteristiche della muscolatura liscia. In alcune malattie (forma catatonica della schizofrenia), le proprietà plastiche dei muscoli scheletrici diventano pronunciate.
Le proprietà fisiologiche dei muscoli includono eccitabilità, conduttività e contrattilità.
Lavoro muscolare. Poiché le estremità del muscolo sono attaccate alle ossa, i punti della sua origine e attacco durante la contrazione si avvicinano l'uno all'altro e i muscoli stessi eseguono un determinato lavoro. Pertanto, il corpo umano o le sue parti, quando i muscoli corrispondenti si contraggono, cambiano posizione, si muovono, superano la resistenza della gravità o, al contrario, cedono a questa forza. In altri casi, quando i muscoli si contraggono, il corpo viene trattenuto certa posizione senza eseguire il movimento. Sulla base di ciò, viene fatta una distinzione tra superare, cedere e trattenere il lavoro muscolare.
Superare il lavoro muscolare viene eseguito quando la forza di contrazione muscolare cambia la posizione di una parte del corpo, di un arto o di un suo collegamento, con o senza carico, vincendo la forza di resistenza.
Inferioreè detto lavoro in cui la forza muscolare è inferiore all'azione della gravità di una parte del corpo (arto) e del carico da essa sostenuto. Il muscolo lavora, ma non si accorcia, ma, al contrario, si allunga; ad esempio, quando è impossibile sollevare o trattenere un oggetto di massa elevata. Con un grande sforzo muscolare, devi abbassare questo corpo sul pavimento o su un'altra superficie.
Trattenere il lavoro viene eseguito se la forza delle contrazioni muscolari mantiene il corpo o il carico in una determinata posizione senza muoversi nello spazio. Ad esempio, una persona sta in piedi o si siede senza muoversi oppure tiene un carico nella stessa posizione. La forza della contrazione muscolare bilancia il peso del corpo o del carico. In questo caso i muscoli si contraggono senza modificare la loro lunghezza (contrazione isometrica).
Il lavoro di superamento e cedimento, quando la forza delle contrazioni muscolari muove il corpo o sue parti nello spazio, può essere considerato un lavoro dinamico. Il lavoro di trattenimento, in cui non avviene alcun movimento dell'intero corpo o di una parte del corpo, è un lavoro statico.
Le ossa collegate dalle articolazioni fungono da leve quando i muscoli si contraggono. In biomeccanica si distingue una leva del primo tipo, in cui i punti di resistenza e di applicazione della forza muscolare si trovano su lati opposti del fulcro, e una leva di secondo tipo, in cui entrambe le forze sono applicate dallo stesso lato del fulcro. il fulcro, a diverse distanze da esso.
Leva del primo tipo si chiama doppia spalla "leva di equilibrio" Il fulcro si trova tra il punto di applicazione della forza (la forza di contrazione muscolare) e il punto di resistenza (gravità, massa dell'organo). Un esempio di tale leva è la connessione tra la colonna vertebrale e il cranio. L'equilibrio si ottiene a condizione che il momento torcente della forza applicata (il prodotto della forza che agisce sull'osso occipitale per la lunghezza del braccio, che è uguale alla distanza dal fulcro al punto di applicazione della forza) sia pari a il momento torcente di gravità (il prodotto della gravità per la lunghezza del braccio, che è pari alla distanza dal punto di appoggio al punto di applicazione della gravità).
Leva del secondo tipo monospalla. In biomeccanica (a differenza della meccanica) è di due tipi. Il tipo di tale leva dipende dalla posizione del punto di applicazione della forza e del punto di azione della gravità, che in entrambi i casi si trovano sullo stesso lato del fulcro. Il primo tipo di leva del secondo tipo (leva di forza) si verifica quando il braccio di applicazione della forza muscolare è più lungo del braccio di resistenza (gravità). Considerando il piede come esempio, possiamo vedere che il fulcro (asse di rotazione) sono le teste delle ossa metatarsali, e il punto di applicazione della forza muscolare (tricipite surale) è calcagno. Il punto di resistenza (peso del corpo) si trova nel punto di articolazione delle ossa della parte inferiore della gamba con il piede (articolazione della caviglia). In questa leva si nota un aumento di forza (il braccio di applicazione della forza è più lungo) e una perdita di velocità di movimento del punto di resistenza (il suo braccio è più corto). Il secondo tipo ha una leva a braccio singolo (leva della velocità) La spalla di applicazione della forza muscolare è più corta della spalla di resistenza, dove viene applicata la forza opposta, la gravità. Per superare la forza di gravità, il cui punto di applicazione si trova a notevole distanza dal punto di rotazione dell'articolazione del gomito (fulcro), è necessaria una forza notevolmente maggiore da parte dei muscoli flessori attaccati vicino all'articolazione del gomito (a il punto di applicazione della forza). In questo caso si ottiene un aumento della velocità e dell'ampiezza di movimento della leva più lunga (punto di resistenza) e una perdita della forza che agisce nel punto di applicazione di tale forza.
forza muscolare
La forza è il prodotto della massa per l'accelerazione ad essa impartita. Quando si eseguono determinati movimenti lavorativi e sportivi, la massima forza muscolare si ottiene sia a causa del maggiore aumento della massa del carico da sollevare o spostare, sia a causa di un aumento dell'accelerazione, cioè una variazione della velocità fino al valore massimo. Nel primo caso aumenta la tensione muscolare e nel secondo aumenta la velocità della sua contrazione. Il movimento negli esseri umani avviene solitamente attraverso una combinazione di contrazione e tensione muscolare. Pertanto all’aumentare della velocità di contrazione aumenta proporzionalmente anche la tensione. Maggiore è la massa del carico, minore è l'accelerazione impartitagli da una persona.
La forza massima di un muscolo viene misurata determinando il carico massimo che può muovere. In tali condizioni isometriche, il muscolo quasi non si contrae e la sua tensione è estrema. Pertanto, il grado di tensione muscolare è espressione della sua forza.
I movimenti di potenza sono caratterizzati dalla massima tensione con un aumento della massa del carico e una velocità costante del suo movimento.
La forza di un muscolo non dipende dalla sua lunghezza, ma dipende principalmente dal suo spessore, dal suo diametro fisiologico, cioè dal numero di fibre muscolari per area della sezione trasversale maggiore. L'area della sezione trasversale fisiologica è l'area della sezione trasversale di tutte le fibre muscolari. Nei muscoli pennati e semipennati questo diametro è maggiore di quello anatomico. Nei muscoli fusiformi e paralleli il diametro fisiologico coincide con quello anatomico. Pertanto i più forti sono i muscoli pennati, poi i semipennati, fusiformi e, infine, i muscoli più deboli con fibre parallele. La forza di un muscolo dipende anche dal suo stato funzionale, dalle condizioni del suo lavoro, dalla frequenza e grandezza massima, dalla somma spaziale e temporale degli impulsi nervosi che gli affluiscono causandone la contrazione, dal numero di unità neuromotorie funzionanti e da impulsi che regolano il metabolismo. La forza muscolare aumenta con l'allenamento e diminuisce con il digiuno e l'affaticamento. Inizialmente aumenta con l’età, poi diminuisce con la vecchiaia.
La forza di un muscolo nella sua massima tensione, sviluppata nella sua massima eccitazione e nella lunghezza più favorevole prima dell'inizio della sua tensione, è chiamata assoluto.
La forza muscolare assoluta viene misurata in chilogrammi o newton (N). La massima tensione muscolare in una persona è causata dallo sforzo volontario.
Parente La forza muscolare viene calcolata come segue. Dopo aver determinato la forza assoluta in chilogrammi o newton, dividerla per il numero di centimetri quadrati della sezione trasversale del muscolo. Ciò consente di confrontare la forza di diversi muscoli dello stesso corpo, la forza dei muscoli con lo stesso nome organismi diversi, così come i cambiamenti nella forza dello stesso muscolo di un dato organismo a seconda dei cambiamenti nel suo stato funzionale. Forza relativa del muscolo scheletrico della rana 2-3 kg, estensore collo umano- 9 kg, muscolo masticatorio - 10 kg, muscolo bicipite brachiale - 11 kg, muscolo tricipite brachiale - 17 kg.
La dinamometria è un metodo per misurare la forza di contrazione di vari gruppi muscolari.
Per la dinamometria ci sono vari tipi dinamometri. Il più comune è un dinamometro a molla (Fig. 1).
Il soggetto lo stringe con la mano tesa. La forza di compressione è indicata da una freccia su una scala speciale. Un'altra modifica è il dinamometro Sternberg (Fig. 2), che ha due ampie maniglie parallele, che il soggetto stringe anche con la mano.
Esistono dinamometri a mercurio (Fig. 3), in cui la forza di pressione sul sensore viene determinata utilizzando un manometro a mercurio.
Un tipo di dinamometria, la dinamografia, è un metodo che consente di registrare graficamente la forza di contrazione muscolare sotto forma di una serie di curve. Questo metodo riflette la dinamica dello sforzo muscolare a lungo termine di un determinato gruppo muscolare.
La dinamometria è utilizzata in antropologia, antropometria, neuropatologia, balneologia, ecc.
20. Affaticamento muscolare
La fatica è una diminuzione o perdita temporanea delle prestazioni, cioè il risultato del lavoro precedente. L'affaticamento di un muscolo nel corpo in condizioni di circolazione sanguigna non dipende solo dalla quantità di muscolo da esso prodotto. lungo lavoro, ma dal numero di onde di eccitazione che arrivano ad esso provocandone la contrazione. A parità di frequenza di irritazione e di altre condizioni uguali, l'affaticamento appare prima con un maggiore carico muscolare. A parità di carico e in altre condizioni uguali, la fatica si manifesta prima con irritazioni più frequenti. All'inizio del lavoro, l'altezza delle contrazioni aumenta, quindi i segni di sviluppo dell'affaticamento sono una graduale diminuzione dell'altezza delle contrazioni, un aumento della loro durata e un aumento delle contratture. Lo sviluppo della fatica dipende dai cambiamenti nel metabolismo, nella circolazione sanguigna, nella temperatura e in altre condizioni. Maggiore è il metabolismo e migliore circolazione sanguigna, subentra la stanchezza successiva. Si verifica molto prima quando il muscolo si contrae, allungato da un carico durante la contrazione isometrica, e successivamente quando si contrae senza carico, e quindi senza tensione.
Se un muscolo viene portato al punto di completo affaticamento a causa dell'irritazione con una corrente elettrica, dopo aver cambiato la direzione della corrente le sue prestazioni vengono immediatamente ripristinate. Questo recupero è spiegato dai cambiamenti nello stato delle proteine muscolari e dagli spostamenti degli ioni ai poli della corrente. Un muscolo isolato riduce il suo lavoro o addirittura smette di contrarsi quando la riserva di glicogeno è pari alla metà della quantità originale. Questi fatti non supportano la teoria dell'esaurimento (Schiff, 1868), che spiega l'affaticamento muscolare con il consumo di sostanze che liberano energia per il suo lavoro. Tuttavia, le riserve di glicogeno nel corpo umano sono limitate e ammontano a 300-400 g, con un lavoro molto intenso vengono consumate in 1,5-2 ore, il che porta a una tale diminuzione della glicemia che il lavoro diventa impossibile. L'introduzione di zucchero nel corpo ripristina le sue prestazioni.
La teoria dell'avvelenamento muscolare durante l'affaticamento dovuto ad un veleno speciale che si accumula in esso - la kenotossina (Weichardt, 1904) si è rivelata infondata. Ma ci sono prove che la fatica è talvolta associata all'avvelenamento delle strutture eccitate da parte di prodotti metabolici, principalmente acidi fosforici e lattici al momento della loro formazione. I prodotti metabolici residui sembrano intasare il corpo e causare affaticamento: la teoria dell'intasamento (Pfluger, 1872).
L'accumulo di acido fosforico e lattico riduce le prestazioni muscolari. Una fibra muscolare isolata, a differenza di un muscolo intero, si stanca molto più tardi con lo stesso numero di impulsi irritanti. Questo è spiegato da prodotti finali il metabolismo viene rimosso più velocemente. In un muscolo allenato, a causa della grande accelerazione dell'analisi e della sintesi delle sostanze che ne garantiscono il funzionamento, l'affaticamento si manifesta più tardi. Dopo il lavaggio vasi sanguigni un muscolo isolato portato al punto di completo affaticamento, quindi, dopo aver eliminato da esso parte dei prodotti metabolici residui, ricomincia a contrarsi nonostante non sia stato ripristinato l'apporto di carboidrati e ossigeno. Questi fatti dimostrano che i prodotti di decadimento residui delle sostanze formate nel muscolo che lavora sono una delle ragioni del suo affaticamento.
Esiste anche una teoria del soffocamento (M. Verworn, 1903), che attribuisce il ruolo principale nella fatica alla mancanza di ossigeno. È noto che il lavoro può durare decine di minuti e anche ore senza fatica, quando il livello di consumo di ossigeno è inferiore al limite di apporto di ossigeno possibile per il lavoratore (vero stato stazionario). Quando il consumo di ossigeno raggiunge il massimo, può essere a un livello costante, ma non soddisfa il fabbisogno di ossigeno del corpo (uno stato stazionario apparente, o meridionale) e il lavoro in questo caso non può durare più di 10-40 minuti.
La fatica è un normale processo fisiologico che porta alla cessazione del lavoro. Durante le pause dal lavoro, le prestazioni muscolari vengono ripristinate. Pertanto, la validità del coinvolgimento degli acidi fini e fosforici nell'insorgenza della fatica non ci consente di trarre l'assurda conclusione che il lavoro sia dannoso, poiché presumibilmente porta all'avvelenamento. È impossibile equiparare l'affaticamento di un muscolo isolato all'affaticamento dell'intero organismo, in cui l'inizio dell'affaticamento dipende da cambiamenti nelle funzioni del sistema nervoso e delle ghiandole endocrine e da cambiamenti nella regolazione del metabolismo, della circolazione sanguigna e respirazione da parte del sistema nervoso centrale. Lo sviluppo dell'affaticamento dipende da una diminuzione delle prestazioni del sistema circolatorio, in particolare del cuore, e del sistema respiratorio.
IN condizioni normali per molto tempo lavoro fisico l'eccitazione e la contrazione dei muscoli sono due processi correlati che si verificano quando si consuma ossigeno, poiché vengono eseguiti attraverso processi chimici molto complessi che culminano nell'ossidazione dei prodotti metabolici residui. Le prestazioni muscolari dopo la fatica vengono ripristinate grazie all'ossidazione di questi prodotti. Pertanto, il consumo di ossigeno durante il lavoro muscolare aumenta in modo significativo. Se l'ossigeno fornito non è sufficiente, durante il lavoro muscolare intenso si verifica una mancanza di ossigeno: un debito di ossigeno. In condizioni di insufficiente ossigeno durante il lavoro, le funzioni del sistema nervoso diminuiscono, causa principale dell'affaticamento. Il debito di ossigeno viene ripagato grazie all'aumento della circolazione sanguigna e della respirazione, non solo durante il lavoro, ma anche al termine dello stesso. Questo rimborso del debito di ossigeno termina solo dopo la completa ossidazione dei prodotti metabolici residui formati durante il lavoro e il completo completamento dei processi di riduzione.
Nella preparazione neuromuscolare, l'affaticamento si sviluppa nella regione della giunzione mioneurale. La teoria fondamentale della fatica, che attribuisce il ruolo principale al suo sviluppo nel sistema nervoso centrale dell'intero organismo, è stata formulata da I, M, Sechenov (1902).
Esistono ampie prove del ruolo principale del sistema nervoso centrale nello sviluppo della fatica. La stanchezza si manifesta quando esposti a stimoli condizionati. Quando si è stanchi, aumenta l'inibizione dei riflessi condizionati e incondizionati. Lo sviluppo della fatica è influenzato dall'afflusso di impulsi afferenti; nel cervello, le emozioni. L’attività muscolare cosciente e volontaria è più faticosa dell’attività automatica e involontaria. Essenziale per l'insorgenza dell'affaticamento è lo stato funzionale del cervello, che cambia: con ipossiemia, ipoglicemia, ipertermia, accumulo di metaboliti nel sangue, cambiamenti nelle funzioni degli organi interni, in particolare del sistema cardiovascolare e respiratorio.
La legge dei carichi medi e dei tassi medi di contrazione è di grande importanza per il lavoro e lo sport.
L'attività fisica provoca un complesso di cambiamenti somatovegetativi nel corpo: aumento della frequenza cardiaca, della gittata sistolica, della pressione sanguigna, del consumo di O2 da parte del corpo, della frequenza respiratoria, ecc. attività fisica il metabolismo segue la via aerobica.
Il duro lavoro è accompagnato dall'attivazione dell'ossidazione anaerobica, a seguito della quale l'acido lattico si accumula nei muscoli e si sviluppa l'affaticamento muscolare.
Fatica - stato fisiologico una persona, derivante da un lavoro duro o a lungo termine, che si esprime in una diminuzione temporanea delle prestazioni.
L'affaticamento muscolare (fisico) e centrale (neuro-mentale) sono solitamente combinati.
L'affaticamento è caratterizzato da una diminuzione della forza e della resistenza muscolare, da una ridotta coordinazione dei movimenti, da un indebolimento della RAM, dall'attenzione e da una diminuzione della velocità di elaborazione delle informazioni. Si presume che le cause dell'affaticamento possano essere l'esaurimento dei depositi di glicogeno e l'indebolimento del processo di risintesi dell'ATP, accumulo cibi acidi metabolismo, deplezione di calcio e affaticamento centri nervosi regolazione delle contrazioni dei singoli gruppi muscolari. Soggettivamente la stanchezza si manifesta sotto forma di stanchezza e bisogno di sonno.
Il riposo è uno stato di riposo o un tipo di attività appositamente organizzata che riduce l'affaticamento e riporta gradualmente le funzioni del corpo alla normalità.
LORO. Sechenov ha stabilito che il lavoro di alcuni gruppi muscolari degli arti elimina l'affaticamento di altri gruppi associati al loro lavoro. Questa disposizione ha costituito la base per la definizione di 2 tipi di riposo: passivo e attivo. Il primo implica il riposo relativo, il secondo l'esecuzione di un tipo di lavoro significativamente diverso dal lavoro normalmente svolto.
Il principale elemento morfo-funzionale dell'apparato neuromuscolare dei muscoli scheletrici è l'unità motoria (UM). Comprende il motoneurone del midollo spinale con le fibre muscolari innervate dal suo assone. All'interno del muscolo, questo assone forma diversi rami terminali. Ciascuno di questi rami forma un contatto: una sinapsi neuromuscolare su una fibra muscolare separata. Gli impulsi nervosi provenienti da un motoneurone causano le contrazioni di un gruppo specifico di fibre muscolari. Le unità motorie di piccoli muscoli che eseguono movimenti fini (muscoli dell'occhio, della mano) contengono un piccolo numero di fibre muscolari. In quelli grandi ce ne sono centinaia di volte di più. Tutte le MU, a seconda delle loro caratteristiche funzionali, sono divise in 3 gruppi:
I. Lento e instancabile. Sono formati da fibre muscolari “rosse”, che hanno meno miofibrille. La velocità di contrazione e la forza di queste fibre sono relativamente piccole, ma non si affaticano facilmente. Pertanto, sono classificati come tonici. La regolazione delle contrazioni di tali fibre viene effettuata da un piccolo numero di motoneuroni, i cui assoni hanno pochi rami terminali. Un esempio è il muscolo soleo.
IIB. Veloce, facilmente stanco. Le fibre muscolari contengono molte miofibrille e sono chiamate "bianche". Si contraggono rapidamente e sviluppano una grande forza, ma si stancano rapidamente. Ecco perché sono chiamati fase uno. I motoneuroni di queste unità motorie sono i più grandi e hanno un assone spesso con numerosi rami terminali. Generano impulsi nervosi ad alta frequenza. Muscoli dell'occhio.
IIA. Veloce, resistente alla fatica. Occupano una posizione intermedia.
22Meccanismo di contrazione muscolare
Il muscolo scheletrico è sistema complesso, convertendo l'energia chimica in lavoro meccanico e calore. Attualmente, i meccanismi molecolari di questa trasformazione sono ben studiati.
Organizzazione strutturale della fibra muscolare. La fibra muscolare è una struttura multinucleare circondata da una membrana e contenente un apparato contrattile specializzato: le miofibrille. Inoltre, i componenti più importanti della fibra muscolare sono i mitocondri, un sistema di tubi longitudinali - il reticolo sarcoplasmatico (reticolo) e un sistema di tubi trasversali - il sistema T. L'unità funzionale dell'apparato contrattile di una cellula muscolare è il sarcomero (Fig. 2.20, A); La miofibrilla è costituita da sarcomeri. I sarcomeri sono separati tra loro da placche Z. I sarcomeri nella miofibrilla sono disposti in sequenza, quindi la contrazione dei sarcomeri provoca la contrazione della miofibrilla e l'accorciamento complessivo della fibra muscolare.
Lo studio della struttura delle fibre muscolari al microscopio ottico ha rivelato le loro striature trasversali. Studi al microscopio elettronico hanno dimostrato che le striature trasversali sono dovute alla speciale organizzazione delle proteine contrattili delle miofibrille: actina (peso molecolare 42.000) e miosina (peso molecolare circa 500.000). I filamenti di actina sono rappresentati da un doppio filamento attorcigliato a doppia elica con un passo di circa 36,5 nm. Questi filamenti sono lunghi 1 µm e hanno un diametro di 6-8 nm, il cui numero raggiunge circa 2000, e sono attaccati ad un'estremità alla piastra Z. Le molecole filamentose della proteina tropomiosina si trovano nelle scanalature longitudinali dell'elica di actina. Con incrementi di 40 nm, una molecola di un'altra proteina, la troponina, è attaccata alla molecola di tropomiosina. La troponina e la tropomiosina svolgono un ruolo importante nei meccanismi di interazione tra actina e miosina. Al centro del sarcomero, tra i filamenti di actina, ci sono spessi filamenti di miosina lunghi circa 1,6 µm. In un microscopio polarizzatore, quest'area è visibile come una striscia di colore scuro (a causa della birifrangenza) - un disco A anisotropico. Al centro è visibile una striscia più chiara H. A riposo non ci sono filamenti di actina al suo interno. Su entrambi i lati del disco A sono visibili strisce isotrope leggere: dischi I formati da filamenti di actina. A riposo, i filamenti di actina e miosina si sovrappongono leggermente in modo che la lunghezza totale del sarcomero sia di circa 2,5 μm. La microscopia elettronica ha rivelato una linea M al centro della banda H, una struttura che contiene i filamenti di miosina. Su una sezione trasversale di una fibra muscolare si può vedere l'organizzazione esagonale del miofilamento: ciascun filo di miosina è circondato da sei fili di actina (Fig. 2.20, B).
La microscopia elettronica mostra che ai lati del filamento di miosina ci sono delle sporgenze chiamate ponti trasversali. Sono orientati rispetto all'asse del filamento di miosina con un angolo di 120°. Secondo i concetti moderni, il ponte trasversale è costituito da una testa e un collo. La testa acquisisce una pronunciata attività ATPasi legandosi all'actina. Il collo ha proprietà elastiche ed è un'articolazione incernierata, quindi la testa del ponte trasversale può ruotare attorno al proprio asse.
L'uso della tecnologia dei microelettrodi in combinazione con la microscopia ad interferenza ha permesso di stabilire che l'applicazione della stimolazione elettrica alla regione della placca Z porta ad una contrazione del sarcomero, mentre la dimensione della zona del disco A non cambia, e la dimensione del le strisce H e I diminuiscono. Queste osservazioni hanno indicato che la lunghezza dei filamenti di miosina non cambia. Risultati simili sono stati ottenuti quando il muscolo veniva allungato: la lunghezza intrinseca dei filamenti di actina e miosina non è cambiata. Come risultato di questi esperimenti, è diventato chiaro che l'area di reciproca sovrapposizione dei filamenti di actina e miosina è cambiata. Questi fatti hanno permesso a N. Huxley e A. Huxley di proporre indipendentemente la teoria dello scorrimento del filo per spiegare il meccanismo della contrazione muscolare. Secondo questa teoria, durante la contrazione, la dimensione del sarcomero diminuisce a causa del movimento attivo dei filamenti sottili di actina rispetto ai filamenti spessi di miosina. Attualmente molti dettagli di questo meccanismo sono stati chiariti e la teoria ha ricevuto conferma sperimentale.
Il meccanismo della contrazione muscolare. Durante il processo di contrazione della fibra muscolare, si verificano le seguenti trasformazioni:
A. Conversione elettrochimica:
1. Generazione del PD.
2. Distribuzione della PD attraverso il sistema T.
3. Stimolazione elettrica della zona di contatto del sistema T e del reticolo sarcoplasmatico, attivazione degli enzimi, formazione di inositolo trifosfato, aumento della concentrazione intracellulare di ioni Ca2+.
B. Trasformazione chemiomeccanica:
4. Interazione degli ioni Ca2+ con la troponina, rilascio di centri attivi sui filamenti di actina.
5. Interazione della testa della miosina con l'actina, rotazione della testa e sviluppo della trazione elastica.
6. Scivolamento dei filamenti di actina e miosina l'uno rispetto all'altro, riducendo le dimensioni del sarcomero, sviluppando tensione o accorciamento della fibra muscolare.
Il trasferimento dell'eccitazione dal motoneurone alla fibra muscolare avviene con l'aiuto del mediatore acetilcolina (ACh). L'interazione dell'ACh con il recettore colinergico della placca terminale porta all'attivazione dei canali sensibili all'ACh e alla comparsa di un potenziale della placca terminale, che può raggiungere i 60 mV. In questo caso, l'area della placca terminale diventa una fonte di corrente irritante per la membrana della fibra muscolare e nelle aree membrana cellulare, adiacente alla piastra terminale, si forma un AP, che si propaga in entrambe le direzioni ad una velocità di circa 3-5 m/s ad una temperatura di 36 oC. Pertanto, la generazione della PD è la prima fase della contrazione muscolare.
La seconda fase è la propagazione della PD nella fibra muscolare attraverso il sistema trasversale dei tubuli, che funge da collegamento tra la membrana superficiale e l'apparato contrattile della fibra muscolare. Il sistema T è in stretto contatto con le cisterne terminali del reticolo sarcoplasmatico di due sarcomeri vicini. La stimolazione elettrica del sito di contatto porta all'attivazione degli enzimi situati nel sito di contatto e alla formazione di inositolo trifosfato. Si attiva l'inositolo trifosfato canali del calcio membrane dei serbatoi terminali, che porta al rilascio di ioni Ca2+ dai serbatoi e ad un aumento della concentrazione intracellulare di Ca2+ da 107 a 105 M. L'insieme dei processi che portano ad un aumento della concentrazione intracellulare di Ca2+ costituisce l'essenza di la terza fase della contrazione muscolare. Pertanto, nelle prime fasi, il segnale elettrico dell'AP viene convertito in un segnale chimico - un aumento della concentrazione intracellulare di Ca2+, cioè una trasformazione elettrochimica.
Con un aumento della concentrazione intracellulare di ioni Ca2+, la tropomiosina si sposta nel solco tra i filamenti di actina e si aprono aree sui filamenti di actina con cui i ponti trasversali della miosina possono interagire. Questo spostamento della tropomiosina è dovuto ad un cambiamento nella conformazione della molecola proteica della troponina in seguito al legame del Ca2+. Di conseguenza, la partecipazione degli ioni Ca2+ al meccanismo di interazione tra actina e miosina è mediata attraverso la troponina e la tropomiosina.
Il ruolo essenziale del calcio nel meccanismo della contrazione muscolare è stato dimostrato in esperimenti utilizzando la proteina aequorina, che emette luce quando interagisce con il calcio. Dopo l'iniezione di equorina, la fibra muscolare è stata sottoposta a stimolazione elettrica e contemporaneamente sono state misurate la tensione muscolare isometrica e la luminescenza dell'equorina. Entrambe le curve erano completamente correlate tra loro (Fig. 2.21). Pertanto, la quarta fase dell'accoppiamento elettromeccanico è l'interazione del calcio con la troponina.
Il quinto stadio successivo dell'accoppiamento elettromeccanico è l'attacco della testa del ponte trasversale al filamento di actina al primo di numerosi centri stabili posizionati in sequenza. In questo caso, la testa della miosina ruota attorno al proprio asse, poiché presenta diversi centri attivi che interagiscono in sequenza con i centri corrispondenti sul filamento di actina. La rotazione della testa porta ad un aumento della trazione elastica del collo del ponte crociato e ad un aumento della tensione. In ogni momento specifico dello sviluppo della contrazione, una parte delle teste dei ponti trasversali è in connessione con il filamento di actina, l'altra è libera, cioè c'è una sequenza della loro interazione con il filamento di actina. Ciò garantisce un processo di riduzione regolare. Nella quarta e quinta fase avviene una trasformazione chemiomeccanica.
La reazione sequenziale di connessione e separazione delle teste dei ponti trasversali con il filamento di actina porta allo scorrimento dei filamenti sottili e spessi l'uno rispetto all'altro e ad una diminuzione delle dimensioni del sarcomero e della lunghezza totale del muscolo, che è la sesta tappa. La totalità dei processi descritti costituisce l'essenza della teoria dello scorrimento del filo
Inizialmente si credeva che gli ioni Ca2+ fungessero da cofattore per l'attività ATPasi della miosina. Ulteriori ricerche hanno confutato questa ipotesi. Nel muscolo a riposo, actina e miosina non hanno praticamente alcuna attività ATPasi. L'attaccamento della testa della miosina all'actina fa sì che la testa acquisisca l'attività dell'ATPasi.
L'idrolisi dell'ATP nel centro ATPasi della testa della miosina è accompagnata da un cambiamento nella conformazione di quest'ultima e dal suo trasferimento in un nuovo stato ad alta energia. Il riattacco della testa della miosina a un nuovo centro sul filamento di actina porta nuovamente alla rotazione della testa, che è fornita dall'energia immagazzinata in essa. In ogni ciclo di connessione e separazione della testa della miosina con l'actina, una molecola di ATP viene scissa per ponte. La velocità di rotazione è determinata dalla velocità di degradazione dell'ATP. È chiaro che le fibre fasiche veloci consumano significativamente più ATP per unità di tempo e trattengono meno energia chimica durante l’esercizio tonico rispetto alle fibre lente. Pertanto, nel processo di trasformazione chemiomeccanica, l'ATP fornisce la separazione della testa di miosina e del filamento di actina e fornisce energia per l'ulteriore interazione della testa di miosina con un'altra parte del filamento di actina. Queste reazioni sono possibili a concentrazioni di calcio superiori a 106M.
I meccanismi descritti di accorciamento delle fibre muscolari suggeriscono che il rilassamento richiede innanzitutto una diminuzione della concentrazione di ioni Ca2+. È stato sperimentalmente dimostrato che il reticolo sarcoplasmatico ha un meccanismo speciale: una pompa del calcio, che restituisce attivamente il calcio ai serbatoi. L'attivazione della pompa del calcio viene effettuata dal fosfato inorganico, che si forma durante l'idrolisi dell'ATP, e la fornitura di energia per il funzionamento della pompa del calcio è dovuta anche all'energia generata durante l'idrolisi dell'ATP. Quindi l'ATP è il secondo il fattore più importante, assolutamente necessario per il processo di rilassamento. Per qualche tempo dopo la morte, i muscoli rimangono morbidi a causa della cessazione dell'influenza tonica dei motoneuroni (vedi Capitolo 4). Quindi la concentrazione di ATP diminuisce al di sotto del livello critico e scompare la possibilità di separazione della testa della miosina dal filamento di actina. Il fenomeno del rigor mortis si verifica con pronunciata rigidità dei muscoli scheletrici.
Modalità di lavoro muscolare
Il lavoro meccanico (A) compiuto da un muscolo è misurato dal prodotto del peso sollevato (P) e della distanza (h): A = P * h kgm. Quando si registra il lavoro di un muscolo di rana isolato, è chiaro che maggiore è il carico, minore è l'altezza alla quale il muscolo lo solleva. Esistono 3 modalità di lavoro muscolare: isotonica, isometrica e auxotonica.
Il regime isotonico (regime di tono muscolare costante) si osserva in assenza di carico sul muscolo, quando il muscolo è fissato ad un'estremità e si contrae liberamente. La tensione al suo interno non cambia. Ciò si verifica quando viene irritato un muscolo isolato della rana, fissato ad un'estremità su un treppiede. Poiché in queste condizioni P = 0, allora lavoro meccanico anche i muscoli è zero (A = 0). In questa modalità, nel corpo umano funziona solo un muscolo: il muscolo della lingua. (Nella letteratura moderna, il termine modalità isotonica si trova anche in relazione a una tale contrazione di un muscolo sotto carico, in cui, al variare della lunghezza del muscolo, la sua tensione rimane invariata, ma in questo caso il lavoro meccanico del muscolo il muscolo non è uguale al proiettile, cioè fa lavoro esterno) .
La modalità isometrica (modalità a lunghezza muscolare costante) è caratterizzata dalla tensione del muscolo quando è fissato su entrambe le estremità o quando il muscolo non riesce a sollevare troppo carico. In questo caso h = 0 e, di conseguenza, anche il lavoro meccanico è nullo (A = 0). Questa modalità si osserva quando si mantiene una determinata posa e quando si esegue un lavoro statico. In questo caso, i processi di formazione e distruzione dei ponti tra actina e miosina si verificano ancora nella fibra muscolare, cioè l'energia viene spesa per questi processi, ma non vi è alcuna reazione meccanica di movimento dei filamenti di actina lungo la miosina. Le caratteristiche fisiologiche di tale lavoro consistono nella valutazione dell'entità del carico e della durata del lavoro.
La modalità auxotonica (modalità mista) è caratterizzata da un cambiamento nella lunghezza e nel tono del muscolo, la cui contrazione provoca lo spostamento del carico. In questo caso viene eseguito il lavoro meccanico del muscolo (A = P? h). Questa modalità si manifesta quando si esegue un lavoro muscolare dinamico anche in assenza di carico esterno, poiché i muscoli superano la forza di gravità che agisce sul corpo umano. Esistono 2 tipi di questa modalità di lavoro muscolare: modalità superante (concentrica) e inferiore (eccentrica).
I vertebrati e gli esseri umani hanno tre tipi di muscoli: muscoli striati dello scheletro, muscoli striati del cuore - miocardio e muscoli lisci, che formano le pareti degli organi interni cavi e dei vasi sanguigni.
L'unità anatomica e funzionale del muscolo scheletrico è unità neuromotoria - motoneurone e l'insieme delle fibre muscolari da esso innervate. Gli impulsi inviati dal motoneurone attivano tutte le fibre muscolari che lo compongono.
I muscoli scheletrici sono costituiti grande quantità fibre muscolari. La fibra del muscolo striato ha forma allungata, il suo diametro va da 10 a 100 micron, la lunghezza della fibra va da diversi centimetri a 10-12 cm La cellula muscolare è circondata da una sottile membrana - il sarcolemma, contiene sarcoplasma (protoplasma) e numerosi nuclei. La parte contrattile della fibra muscolare sono lunghi filamenti muscolari - miofibrille, costituiti principalmente da actina, che corrono all'interno della fibra da un'estremità all'altra, con striature trasversali. La miosina nelle cellule muscolari lisce è dispersa, ma contiene molte proteine che svolgono un ruolo importante nel mantenimento della contrazione tonica a lungo termine.
Durante il periodo di riposo relativo, i muscoli scheletrici non si rilassano completamente e mantengono un grado moderato di tensione, cioè tono muscolare.
Funzioni principali tessuto muscolare:
motore – garantendo il movimento
statico – garantendo la fissazione, anche in una determinata posizione
recettore – i muscoli hanno recettori che permettono loro di percepire i propri movimenti
stoccaggio: l'acqua e alcuni nutrienti vengono immagazzinati nei muscoli.
Proprietà fisiologiche dei muscoli scheletrici:
Eccitabilità. Inferiore all'eccitabilità tessuto nervoso. L'eccitazione si diffonde lungo la fibra muscolare.
Conduttività. Minore conduttività del tessuto nervoso.
Periodo refrattario il tessuto muscolare dura più a lungo del tessuto nervoso.
Labilità il tessuto muscolare è significativamente inferiore al tessuto nervoso.
Contrattilità– la capacità di una fibra muscolare di modificare la propria lunghezza e il grado di tensione in risposta alla stimolazione di una forza di soglia.
Con la contrazione isotonica, la lunghezza della fibra muscolare cambia senza cambiare il tono. Con la contrazione isometrica, la tensione della fibra muscolare aumenta senza modificarne la lunghezza.
A seconda delle condizioni di stimolazione e dello stato funzionale del muscolo, può verificarsi una singola contrazione continua (tetanica) o una contrattura del muscolo.
Contrazione del singolo muscolo. Quando un muscolo viene irritato da un singolo impulso di corrente, si verifica un'unica contrazione muscolare.
L'ampiezza di una singola contrazione muscolare dipende dal numero di miofibrille che si contraggono in quel momento. L'eccitabilità dei singoli gruppi di fibre è diversa, quindi la forza della corrente di soglia provoca una contrazione solo delle fibre muscolari più eccitabili. L’ampiezza di tale riduzione è minima. All'aumentare dell'intensità della corrente irritante, nel processo di eccitazione vengono coinvolti anche gruppi di fibre muscolari meno eccitabili; l'ampiezza delle contrazioni si riassume e cresce fino a quando nel muscolo non rimangono più fibre che non siano coperte dal processo di eccitazione. In questo caso si registra la massima ampiezza di contrazione, che non aumenta, nonostante un ulteriore aumento dell'intensità della corrente irritante.
Contrazione tetanica. In condizioni naturali, le fibre muscolari non ricevono singoli, ma una serie di impulsi nervosi, ai quali il muscolo risponde con una contrazione prolungata, tetanica o tetanica. Solo i muscoli scheletrici sono capaci di contrazione tetanica. La muscolatura liscia e la muscolatura striata del cuore non sono in grado di contrarsi tetanica a causa di un lungo periodo refrattario.
Il tetano si verifica a causa della somma delle contrazioni dei singoli muscoli. Perché si manifesti il tetano è necessaria l'azione di irritazioni ripetute (o impulsi nervosi) sul muscolo ancor prima che la sua singola contrazione finisca.
Se gli impulsi irritanti sono ravvicinati e ciascuno di essi si verifica nel momento in cui il muscolo ha appena iniziato a rilassarsi, ma non ha ancora avuto il tempo di rilassarsi completamente, si verifica una contrazione di tipo seghettato (tetano dentato).
Se gli impulsi irritanti sono così ravvicinati che ciascuno successivo avviene in un momento in cui il muscolo non ha ancora avuto il tempo di passare al rilassamento dall'irritazione precedente, cioè avviene al culmine della sua contrazione, allora una contrazione lunga e continua si verifica, chiamato tetano liscio.
Tetano liscio– il normale stato di funzionamento dei muscoli scheletrici è determinato dall’arrivo di impulsi nervosi dal sistema nervoso centrale con una frequenza di 40-50 al secondo.
Tetano seghettato si verifica con una frequenza di impulsi nervosi fino a 30 per 1 s. Se un muscolo riceve 10-20 impulsi nervosi al secondo, allora è in uno stato tono muscolare , cioè. moderato grado di tensione.
Affaticamento muscolare. Con una stimolazione ritmica prolungata nel muscolo, si sviluppa l'affaticamento. I suoi segni sono una diminuzione dell'ampiezza delle contrazioni, un aumento dei loro periodi di latenza, un prolungamento della fase di rilassamento e, infine, l'assenza di contrazioni con continua irritazione.
Un altro tipo di contrazione muscolare prolungata è la contrattura. Continua anche quando lo stimolo viene rimosso. La contrattura muscolare si verifica quando si verifica un disordine metabolico o un cambiamento nelle proprietà delle proteine contrattili del tessuto muscolare. Le cause della contrattura possono essere l'avvelenamento con determinati veleni e farmaci, disturbi metabolici, aumento della temperatura corporea e altri fattori che portano a cambiamenti irreversibili proteine del tessuto muscolare.
Caratteristiche fisiologiche della muscolatura liscia.
I muscoli lisci formano le pareti ( strato muscolare) organi interni e vasi sanguigni. Non sono presenti striature trasversali nelle miofibrille muscolari lisce. Ciò è dovuto alla disposizione caotica delle proteine contrattili. Le fibre muscolari lisce sono relativamente più corte.
Muscolo liscio meno eccitabile rispetto a quelli striati. L'eccitazione si diffonde attraverso di loro a bassa velocità - 2-15 cm/s. L'eccitazione nella muscolatura liscia può essere trasmessa da una fibra all'altra, a differenza delle fibre nervose e delle fibre dei muscoli striati.
La contrazione della muscolatura liscia avviene più lentamente e per un periodo di tempo più lungo.
Il periodo refrattario nei muscoli lisci è più lungo che nei muscoli scheletrici.
Una proprietà importante della muscolatura liscia è la sua grandezza plastica, cioè. la capacità di mantenere la lunghezza data dallo stretching senza modificare la tensione. Questa proprietàè essenziale, poiché alcuni organi addominali (utero, vescica, cistifellea) a volte si allungano notevolmente.
Caratteristica la muscolatura liscia è la loro capacità di operare automaticamente, che è fornito da elementi nervosi incorporati nelle pareti degli organi muscolari lisci.
Uno stimolo adeguato per la muscolatura liscia è il loro allungamento rapido e forte, che è di grande importanza per il funzionamento di molti organi della muscolatura liscia (uretere, intestino e altri organi cavi).
Anche la caratteristica della muscolatura liscia è la loro elevata sensibilità ad alcune sostanze biologicamente attive(acetilcolina, adrenalina, norepinefrina, serotonina, ecc.).
I muscoli lisci sono innervati dal simpatico e dal parasimpatico nervi autonomi, che, di regola, hanno l'effetto opposto sul loro stato funzionale.
Proprietà fondamentali del muscolo cardiaco.
La parete del cuore è composta da 3 strati. Lo strato intermedio (miocardio) è costituito da muscolo striato. Il muscolo cardiaco, come i muscoli scheletrici, ha la proprietà dell'eccitabilità, la capacità di condurre l'eccitazione e la contrattilità. A caratteristiche fisiologiche muscolo cardiaco comprendono un periodo refrattario prolungato e automatismo.
Eccitabilità del muscolo cardiaco. Il muscolo cardiaco è meno eccitabile del muscolo scheletrico. Affinché l'eccitazione si verifichi nel muscolo cardiaco, è necessario uno stimolo più forte rispetto a quello del muscolo scheletrico.
Conduttività. L'eccitazione lungo le fibre del muscolo cardiaco viene effettuata a una velocità inferiore rispetto alle fibre del muscolo scheletrico.
Contrattilità. La reazione del muscolo cardiaco non dipende dalla forza della stimolazione applicata. Il muscolo cardiaco si contrae il più possibile sia alla soglia che alla stimolazione più forte.
Periodo refrattario. Il cuore, a differenza di altri tessuti eccitabili, ha un periodo refrattario significativamente pronunciato ed esteso. È caratterizzato forte calo eccitabilità dei tessuti durante il periodo della sua attività. Per questo motivo, il muscolo cardiaco non è in grado di contrarsi tetanica (a lungo termine) e svolge il suo lavoro come contrazione di un singolo muscolo.
Automaticità del cuore. Fuori dal corpo, in determinate condizioni, il cuore è in grado di contrarsi e rilassarsi, mantenendo il ritmo corretto. Viene chiamata la capacità del cuore di contrarsi ritmicamente sotto l'influenza degli impulsi che sorgono al suo interno automaticità.
Elettromiografia (dal greco mys, myos - muscolo, grafo - scrivo) - registrazione dei potenziali elettrici; muscoli scheletrici. L'elettromiografia viene utilizzata come metodo per studiare la funzione normale e compromessa sistema muscoloscheletrico esseri umani e animali. L'elettromiografia comprende tecniche per studiare l'attività elettrica dei muscoli a riposo, durante le contrazioni volontarie, involontarie e causate da stimoli artificiali.
L'elettromiografia viene utilizzata per studiare lo stato funzionale e caratteristiche funzionali fibre muscolari, unità motorie, trasmissione neuromuscolare, tronchi nervosi, apparato segmentale del midollo spinale, nonché strutture soprasegmentali; Studiano la coordinazione dei movimenti, lo sviluppo delle capacità motorie durante vari tipi di esercizi lavorativi e sportivi, la ristrutturazione del lavoro dei muscoli trapiantati e l'affaticamento. Sulla base dell'elettromiografia, è stato creato un metodo per controllare le biocorrenti muscolari, che ha trovato uso pratico quando si controllano le cosiddette protesi bioelettriche (vedi Protesi).
Un elettromiogramma è una curva ottenuta su carta fotografica, pellicola o carta durante la registrazione dei potenziali elettrici dei muscoli scheletrici. Può essere registrato utilizzando un dispositivo speciale chiamato elettromiografo o altri dispositivi utilizzati per registrare i biopotenziali. Il dispositivo, di regola, ha almeno due canali di registrazione. Ciascun canale include elettrodi di uscita, un amplificatore biopotenziale e un dispositivo di registrazione. La maggior parte degli elettromiografi include un dispositivo per il monitoraggio visivo e uditivo (Figura 1).
Riso. 1. Schema del dispositivo per l'elettromiografia.
La principale fonte di fluttuazioni del potenziale elettrico dei muscoli è il processo di eccitazione che si diffonde lungo le fibre muscolari. Tuttavia, poiché l'elettromiogramma viene registrato nella regione dei punti motori (vedi Elettrodiagnostica), parte del potenziale elettrico è il potenziale che si forma quando le placche terminali vengono eccitate. I potenziali elettrici dei muscoli scheletrici possono essere condotti a livello intracellulare o extracellulare.
La scarica intracellulare dei potenziali elettrici delle singole fibre muscolari nell'uomo consente di determinare quelle caratteristiche che erano state precedentemente studiate negli studi sui microelettrodi su animali o farmaci: l'entità dei potenziali di membrana delle fibre muscolari, la depolarizzazione e l'iperpolarizzazione delle membrane, ecc. (vedi Bioelettrico fenomeni). Numerosi autori chiamano la registrazione dei potenziali intracellulari nei muscoli scheletrici elettromiografia intracellulare.
La rimozione extracellulare dei potenziali elettrici viene effettuata utilizzando due metodi:
1) utilizzando elettrodi con superficie di abduzione relativamente piccola (centesimi di millimetro quadrato), immersi nel muscolo mediante aghi (Fig. 2, 1-3); Inoltre, in tutti i casi, ad eccezione dell'elettrocatetere unipolare, entrambi gli elettrodi dell'elettrocatetere si trovano a breve distanza l'uno dall'altro (solitamente inferiore a 0,5 mm); 2) utilizzando elettrodi con una superficie abduttiva relativamente ampia (30-100 mm2), solitamente posizionati sulla pelle sopra il muscolo a una distanza relativamente grande l'uno dall'altro (1-2 cm) (Fig. 2, 4-6). Nel primo caso è consuetudine parlare di lead “locale”, nel secondo di lead “globale”. La derivazione “locale” consente di studiare i potenziali elettrici che si presentano in un piccolo volume di tessuto muscolare: potenziali delle singole unità motorie, potenziali totali piccola quantità unità motorie, in condizioni patologiche - i potenziali delle singole fibre muscolari. L'oggetto principale di studio è l'unità motoria. Questo concetto originariamente significava un insieme di fibre muscolari innervate da un motoneurone.
La moderna comprensione della struttura e della funzione del sistema nervoso centrale si basa sulla teoria neurale.
Il sistema nervoso è costituito da due tipi di cellule: nervose e gliali, e il numero di queste ultime è 8-9 volte superiore al numero delle cellule nervose. Tuttavia, sono i neuroni che forniscono tutta la varietà di processi associati alla trasmissione e all'elaborazione delle informazioni.
Un neurone, una cellula nervosa, è un'unità strutturale e funzionale del sistema nervoso centrale. I singoli neuroni, a differenza di altre cellule del corpo che agiscono in modo isolato, “lavorano” come una singola unità. Le loro funzioni sono trasmettere informazioni (sotto forma di segnali) da una parte all'altra del sistema nervoso, scambiare informazioni tra il sistema nervoso e aree diverse corpi. In questo caso, i neuroni trasmittenti e riceventi sono combinati in reti e circuiti nervosi.
I processi di elaborazione delle informazioni più complessi avvengono nelle cellule nervose. Con il loro aiuto si formano le risposte del corpo (riflessi) agli stimoli esterni ed interni.
I neuroni hanno una serie di caratteristiche comuni a tutte le cellule del corpo. Indipendentemente dalla sua posizione e dalle sue funzioni, qualsiasi neurone, come qualsiasi altra cellula, ha una membrana plasmatica che definisce i confini della singola cellula. Quando un neurone comunica con altri neuroni o percepisce cambiamenti nell'ambiente locale, lo fa attraverso la membrana e i meccanismi molecolari in essa contenuti. Vale la pena notare che la membrana di un neurone ha una forza significativamente maggiore rispetto ad altre cellule del corpo.
Tutto quello che c'è dentro membrana plasmatica(ad eccezione del nucleo) è chiamato citoplasma. Contiene gli organelli citoplasmatici necessari affinché il neurone esista e svolga il suo lavoro. I mitocondri forniscono energia alla cellula utilizzando zucchero e ossigeno per sintetizzare speciali molecole ad alta energia che la cellula utilizza secondo necessità. I microtubuli - sottili strutture di supporto - aiutano il neurone a mantenere una certa forma. Rete di tubuli della membrana interna attraverso i quali la cellula si distribuisce sostanze chimiche, necessario al suo funzionamento, è chiamato reticolo endoplasmatico.
Esistono due tipi di reticolo endoplasmatico: “ruvido” e “liscio”. Le membrane ruvide (granulari) sono costellate di ribosomi, cellule necessarie per la sintesi delle sostanze proteiche da esso secrete. L'abbondanza di elementi del reticolo “grezzo” nei neuroni li caratterizza come cellule con un'attività molto intensa. Un altro tipo di reticolo plasmatico, quello liscio, detto anche apparato di Golgi, “impacchetta” le sostanze sintetizzate dalla cellula in apposite “sacche” costruite dalle membrane del reticolo liscio. Il compito di questo organello neuronale è trasportare le secrezioni sulla superficie cellulare.
Al centro del citoplasma c'è il nucleo che, come tutte le cellule dotate di nucleo, contiene informazioni genetiche codificate nella struttura chimica dei geni. In base a queste informazioni, una cellula completamente formata sintetizza sostanze specifiche che determinano la forma, la chimica e la funzione di questa cellula. Tuttavia, a differenza della maggior parte delle altre cellule del corpo, i neuroni maturi non possono dividersi. Pertanto, gli elementi chimici geneticamente determinati di qualsiasi neurone devono garantire la conservazione e il cambiamento delle sue funzioni per tutta la sua vita. Nei neuroni di grandi dimensioni, il nucleo costituisce 1/3-1/4 della dimensione del loro corpo. I nucleoli inclusi nella sua composizione sono coinvolti nel fornire alla cellula acidi ribonucleici e proteine (nei motoneuroni, ad esempio, attività motoria i nucleoli animali aumentano significativamente di dimensioni).
Allo stesso tempo, i neuroni, a differenza di altre cellule del corpo, hanno caratteristica essenziale, essi, oltre al corpo (soma), sono dotati di processi: numerosi brevi processi ramificati a forma di albero - i dendriti (tradotto dal greco - albero) servono come una sorta di input neuronali, attraverso i quali i segnali entrano nella cellula nervosa. Hanno una superficie ruvida creata da piccoli ispessimenti: spine, come perline, infilate su un dendrite. Ciò aumenta la superficie del neurone e massimizza la raccolta delle informazioni.
L'uscita di un neurone è un processo lungo e regolare che si estende dal gene - un assone (dal greco asse - asse), che trasmette gli impulsi nervosi ulteriormente ad un'altra cellula nervosa o organo funzionante (Fig. 1). Gli assoni di molti neuroni sono ricoperti da una guaina mielinica. È formato da cellule di Schwann, ripetutamente (fino a 10 o più strati) “avvolte” come un nastro isolante attorno al tronco dell'assone. Tuttavia, gli accoppiamenti delle cellule di Schwann posizionate sull'assone non si toccano. Tra di loro rimangono stretti spazi vuoti: i nodi di Ranvier. Solo qui la fibra nervosa entra in contatto diretto con il fluido extracellulare. Pertanto, nel sistema nervoso dei mammiferi, l'onda di un impulso nervoso che si propaga non scorre uniformemente, ma si sposta bruscamente (saltatoriamente) da un'intercettazione all'altra, il che accelera notevolmente il processo di propagazione dell'impulso.
Per quanto riguarda la parte iniziale dell'assone nel punto in cui esce dal corpo cellulare (la zona della “protuberanza dell'assone”), è priva di guaina mielinica. La membrana di questa parte non mielinizzata del neurone, il cosiddetto segmento iniziale, è altamente eccitabile. Pertanto, viene chiamata zona trigger, poiché è qui che inizia l'eccitazione del neurone.
Non c'è bisogno di dire che anche le connessioni intracerebrali richiedono processi molto lunghi, per non parlare degli assoni che si estendono oltre il sistema nervoso centrale - fino ai muscoli, alle ghiandole e agli organi interni. Raccolti in fasci, formano i nervi.
Se un neurone forma connessioni in uscita con un grande membro di altre cellule, il suo assone può ramificarsi molte volte in modo che i segnali possano raggiungere ciascuno di essi; il numero di tali rami (termopoli) è enorme e varia da 1.000 a 10.000 o più. Inoltre, l'assone è in grado di emettere ulteriori rami - collaterali, lungo i quali l'eccitazione si allontana dal percorso principale. I processi separati dal corpo cellulare non possono esistere a lungo e muoiono. Il corpo cellulare, al contrario, li rigenera. Naturalmente questo vale solo per la parte centrale del processo. A volte i processi di rigenerazione dei processi avvengono a velocità enorme: fino a 30 micron al minuto.
Va notato che è proprio a causa della presenza di processi che i neuroni, come le cellule, sono stati scoperti più tardi rispetto ad altre cellule del corpo umano e animale. Ciò è comprensibile, poiché il neurone con tutti i suoi processi non potrebbe rientrare nel campo visivo del microscopio. Pertanto, inizialmente alle cellule stesse non è stata data la dovuta importanza, considerandole come un addensamento tra tanti processi.
La forma della cellula nervosa, le sue dimensioni e la posizione dei suoi processi sono varie e dipendono dallo scopo funzionale del neurone (Fig. 2).
Ogni singolo neurone è unico e diverso dal suo genere, a differenza delle altre cellule del corpo. La dimensione dei neuroni è molto variabile: quelli più grandi sono decine e centinaia di volte più grandi di quelli più piccoli. Ad esempio, il diametro dei granuli nel cervelletto è di 7,0 micron e quello dei motoneuroni nel midollo spinale è di 70,0.
La densità dei neuroni in alcune parti del sistema nervoso centrale è molto alta. Sì, nella corteccia emisferiè pari a 40.000 cellule per 1 mm3. Nessuno può rispondere esattamente alla domanda su quanti neuroni contenga il cervello degli esseri umani e degli animali altamente organizzati, ma si ritiene che il loro numero sia di circa decine di miliardi.
Fibre nervose non mielinizzate- uno strato di cellule di Schwann, tra di loro ci sono spazi a fessura. La membrana cellulare è in contatto ovunque ambiente. Quando viene applicata l'irritazione, l'eccitazione si verifica nel sito di azione dell'irritante. Le fibre nervose non mielinizzate hanno proprietà elettrogeniche (la capacità di generare impulsi nervosi) per tutta la loro lunghezza.
Fibre nervose mielinizzate- ricoperti da strati di cellule di Schwann, che in alcuni punti formano nodi di Ranvier (aree prive di mielina) ogni 1 mm. La durata del nodo di Ranvier è di 1 µm. La guaina mielinica svolge funzioni trofiche e isolanti (alta resistenza). Le aree ricoperte di mielina non hanno proprietà elettrogeniche. Sono posseduti dai nodi di Ranvier. L'eccitazione avviene nel nodo di Ranvier più vicino al sito d'azione dello stimolo. Ai nodi di Ranvier c'è un'alta densità di canali del Na, quindi ad ogni nodo di Ranvier si verifica un aumento degli impulsi nervosi.
I nodi di Ranvier funzionano come relè (generano e amplificano gli impulsi nervosi).
Il meccanismo di eccitazione lungo la fibra nervosa
1885 - L. Herman - Tra le sezioni eccitate e non eccitate della fibra nervosa si formano correnti circolari.
Quando agisce uno stimolo, esiste una differenza di potenziale tra la superficie esterna e quella interna del tessuto (aree con cariche diverse). Tra queste zone si forma una corrente elettrica (movimento degli ioni Na+). All'interno della fibra nervosa si genera una corrente dal polo positivo al polo negativo, cioè la corrente viene diretta dalla zona eccitata a quella non eccitata. Questa corrente esce attraverso l'area non eccitata e la fa ricaricare. SU superficie esterna La corrente della fibra nervosa fluisce dalla zona non eccitata a quella eccitata. Questa corrente non modifica lo stato della zona eccitata, poiché è in uno stato di refrattarietà.
Prova della presenza di correnti circolari: la fibra nervosa viene posta in una soluzione di NaCl e viene registrata la velocità di eccitazione. Successivamente la fibra nervosa viene immersa nell'olio (la resistenza aumenta) - la velocità di conduzione diminuisce del 30%. Successivamente, la fibra nervosa viene lasciata nell'aria: la velocità di eccitazione viene ridotta del 50%.
Caratteristiche della conduzione dell'eccitazione lungo le fibre nervose mielinizzate e non mielinizzate:
1) fibre mieliniche - hanno una guaina con proprietà elettrogeniche ad alta resistenza solo nei nodi di Ranvier. Sotto l'influenza di uno stimolo, l'eccitazione avviene nel nodo di Ranvier più vicino. L'intercettazione vicina è in uno stato di polarizzazione. La corrente risultante provoca la depolarizzazione dell'intercettazione adiacente. Ai nodi di Ranvier c'è un'alta densità di canali Na, quindi ad ogni nodo successivo si forma un potenziale d'azione leggermente più grande (in ampiezza), per questo l'eccitazione si diffonde senza decremento e può saltare su più nodi. Questa è la teoria saltatoria di Tasaki. Prova della teoria: i farmaci sono stati iniettati nella fibra nervosa, bloccando diverse intercettazioni, ma la conduzione dell'eccitazione è stata registrata anche dopo. Questo è un metodo altamente affidabile e redditizio, poiché si eliminano i danni minori, si aumenta la velocità di eccitazione e si riducono i costi energetici;
2) fibre non mielinizzate: la superficie ha proprietà elettrogeniche ovunque. Pertanto, a una distanza di diversi micrometri si formano piccole correnti circolari. L'eccitazione ha l'aspetto di un'onda in costante movimento. Questo metodo è meno redditizio: maggiori costi energetici (per il funzionamento della pompa Na-K), minore velocità di eccitazione.
Il meccanismo per condurre l'eccitazione lungo le fibre nervose dipende dal loro tipo. Esistono due tipi di fibre nervose: mielinizzate e non mielinizzate.
I processi metabolici nelle fibre non mielinizzate non forniscono una rapida compensazione per il dispendio energetico. La diffusione dell'eccitazione avverrà con graduale attenuazione, con decremento. Il comportamento decrescente dell'eccitazione è caratteristico di un sistema nervoso poco organizzato. L'eccitazione si propaga a causa di piccole correnti circolari che si generano nella fibra o nel liquido circostante. Tra ecc.
Stomaco e intestino, cambiamenti nel tono dei vasi sanguigni, mantenimento del tono della vescica - sono causati dalla contrazione della muscolatura liscia. Il lavoro del cuore è assicurato dalla contrazione dei muscoli cardiaci.
YouTube enciclopedico
1 / 3
Tropomiosina e troponina e loro ruolo nel controllo della contrazione muscolare
Contrazione muscolare (filamenti di actina e miosina)
La struttura di una cellula muscolare
Sottotitoli
Nell'ultima lezione abbiamo imparato come la miosina - o meglio la miosina II, che ha due teste su due catene intrecciate tra loro - come la miosina II interagisce con l'ATP e quindi spinge o scivola lungo i filamenti di actina. All'inizio è attaccata la miosina. L'ATP si lega quindi alla testa della miosina. La miosina si stacca dall'actina. L'ATP viene idrolizzato in ADP e fosfati. Questo rilascia energia. Porta la miosina in uno stato ad alta energia. Grazie a questa energia, la miosina si attacca alla sezione successiva del filamento di actina; il gruppo fosfato viene quindi rilasciato, provocando il cambiamento di forma della proteina. In questo caso l'energia accumulata dalla miosina è diretta ad agire sulle fibre di actina. La miosina si sposta a sinistra o l'actina si sposta a destra, a seconda dell'angolo di visione. Nelle lezioni seguenti parleremo di cosa sono legate l'actina e la miosina. Probabilmente hai un paio di domande. La testa della miosina ha speso così tanta energia per spingere la fibra di actina. E allo stesso tempo probabilmente è stata esposta a resistenza o ad altre forze. Cosa succede quando si disconnette? Nella fase successiva del ciclo, quando l’ATP si legherà nuovamente ad esso, il filamento di actina ritornerà nella sua posizione originale? Soprattutto sotto l'influenza della tensione. Devi capire che l'actina è influenzata non solo da questa testa di miosina. Ce ne sono molti altri. Sono lungo l'intera catena dell'actina. Sia qui che là. E tutti lavorano al proprio ritmo. E quando alcuni di loro sono distaccati, altri sono nella fase del “colpo di potenza”, e altri sono attaccati. Non è che una testa di miosina si stacchi e il filamento di actina ritorni nella sua posizione originale. Un'altra domanda: cosa avvia e cosa ferma questo processo? Dopotutto, controlliamo i nostri muscoli. Quindi cosa innesca il processo di scorrimento della miosina lungo le fibre di actina? Altre due proteine svolgono questo ruolo. Tropomiosina e troponina. Lo scriverò. Tropomiosina. E in un colore diverso: la troponina. Ora rappresenterò schematicamente un filamento di actina. Eccolo. Questo è il mio filamento di actina. In realtà è una struttura a spirale. Aggiungo una seconda riga. Sono come le rientranze di una spirale; la precisione dell’immagine non è importante per noi adesso. Nell'ultima lezione abbiamo esaminato le azioni delle teste di miosina. Subiscono un certo ciclo, nelle diverse fasi del quale ricevono e cedono energia, cambiano forma e ritornano al loro stato originale. Ma esiste anche una proteina associata all'actina: la tropomiosina. Sembra essere avvolto attorno all'actina. Disegnerò. Questa è recitazione. Questa è una delle teste della miosina II. Miosina. Ed ecco la tropomiosina. È avvolto attorno all'actina. Descriverò schematicamente - ecco come viene avvolto - indicherò con una linea tratteggiata la parte invisibile della catena della tropomiosina. Linea continua: visibile e così via. La tropomiosina avvolge la catena dell'actina. La tropomiosina è attaccata all'actina da un'altra proteina chiamata troponina. Diciamo che si allega qui - questo, ovviamente, disegno schematico- qui e qui... usando la troponina. Lo scriverò. La tropomiosina è indicata in verde chiaro. E la troponina è come i chiodi che fissano la tropomiosina all'actina. Quando un muscolo non si contrae, la tropomiosina non produce miosina... In effetti, il processo non è ancora del tutto compreso. La ricerca continua. Ma sembra che la tropomiosina impedisca alla testa della miosina di attaccarsi alla catena di actina e di scivolare lungo di essa; e quando la testa è già attaccata alla catena di actina, la tropomiosina ne impedisce il distacco ed elimina così la possibilità di tale scorrimento. Quindi, la tropomiosina blocca la testa della miosina, blocca la testa della miosina - qui abbiamo la testa della miosina - e le impedisce di scivolare lungo le fibre di actina... non le permette di scivolare lungo le fibre di actina... O blocca fisicamente il sito per legare, oppure - la testa è già attaccata - eliminando la possibilità del suo distacco. In entrambi i casi l'interazione con l'actina è impossibile; Per sbloccare la testa, le troponine devono cambiare forma. E possono cambiare forma solo con un'alta concentrazione di calcio. Se abbiamo abbastanza ioni calcio alta concentrazione, - alcuni di questi ioni si uniranno alla troponina; a causa di ciò cambierà la conformazione, cioè la forma, della troponina. Questo, a sua volta, cambierà la forma della tropomiosina. Lo scriverò. La tropomiosina svolge una funzione bloccante; ma ad un'alta concentrazione di ioni calcio, tali ioni si legano alla troponina, che cambia la conformazione della troponina e “disattiva” la funzione di blocco della tropomiosina. ... “spegne” la funzione bloccante della tropomiosina... Quindi, un'alta concentrazione di ioni calcio, legandosi alla troponina, “spegne” la tropomiosina... E ora si sono verificate le condizioni con cui abbiamo iniziato l'ultima lezione: la miosina e l'actina possono influenzarsi a vicenda: spingere, scivolare... possono essere chiamate diversamente. E se la concentrazione di calcio diminuisce (bassa concentrazione di ioni calcio), tali ioni vengono rilasciati dalla troponina. Se gli ioni calcio scarseggiano, gli ioni trattenuti dalla troponina vengono rilasciati. Di conseguenza la troponina ritorna alla sua conformazione originaria. ...la troponina ritorna... E la tropomiosina ricomincia a bloccare la miosina. ...permette alla tropomiosina di bloccare nuovamente la miosina... Naturalmente, questo meccanismo non può essere definito semplice. È stato scoperto di recente, 50-60 anni fa. Immagina quanti esperimenti sono già stati effettuati e saranno ancora necessari... Ma lo schema in sé non è complicato. Quando c'è carenza di calcio, la tropomiosina blocca la capacità della miosina di legarsi e interagire con la catena dell'actina. Ad una concentrazione sufficientemente elevata di calcio, i suoi ioni si legano alla troponina, che ancora la tropomiosina alla catena dell'actina, gli ioni calcio cambiano la conformazione della troponina, la tropomiosina si stacca e la miosina può fare il suo lavoro. Puoi immaginare schematicamente il meccanismo di contrazione muscolare e il controllo di tale contrazione. Se c'è un'alta concentrazione di calcio nella cellula, il muscolo si contrarrà. Se la concentrazione calcio basso, la troponina rilascerà ioni, la miosina verrà bloccata e il muscolo si rilasserà. Sottotitoli a cura della comunità Amara.org
introduzione
La base di tutti i tipi di contrazione muscolare è l'interazione tra actina e miosina. Nei muscoli scheletrici, le miofibrille (circa due terzi del peso secco del muscolo) sono responsabili della contrazione. Le miofibrille sono strutture spesse 1 - 2 micron, costituite da sarcomeri - strutture lunghe circa 2,5 micron, costituite da filamenti di actina e miosina (sottili e spessi) e dischi Z collegati ai filamenti di actina. La contrazione avviene con un aumento della concentrazione di ioni Ca 2+ nel citoplasma a seguito dello scorrimento dei filamenti di miosina rispetto ai filamenti di actina. La fonte dell’energia di contrazione è l’ATP. L'efficienza di una cellula muscolare è di circa il 50%, il muscolo nel suo insieme non supera il 20%. La massima forza muscolare non viene raggiunta condizioni reali; Non tutte le cellule muscolari vengono utilizzate contemporaneamente e si contraggono con la massima forza, altrimenti, quando si contraggono molti muscoli scheletrici, i tendini o le ossa verranno danneggiati (cosa che a volte si osserva con forti crampi). L'efficienza muscolare dipende anche dalle condizioni esterne; ad esempio con il freddo diminuisce notevolmente, poiché è più importante per l'organismo mantenere la temperatura corporea.
Scorrimento della miosina rispetto all'actina
Le teste di miosina scompongono l'ATP e, a causa dell'energia rilasciata, cambiano conformazione, scorrendo lungo i filamenti di actina. Il ciclo può essere suddiviso in 4 fasi:
- La testa libera della miosina si lega all'ATP e lo idrolizza in ADP e fosfato e rimane associata ad essi. (Un processo reversibile: l'energia rilasciata a seguito dell'idrolisi viene immagazzinata nella mutata conformazione della miosina).
- Le teste si legano debolmente alla successiva subunità di actina, il fosfato viene rilasciato e questo porta ad un forte legame della testa di miosina al filamento di actina. Questa reazione è già irreversibile.
- La testa subisce un cambiamento conformazionale che tira il filamento spesso verso il disco Z (o, equivalentemente, le estremità libere dei filamenti sottili l'una verso l'altra).
- Viene rilasciato ADP, per questo la testa viene separata dal filamento di actina. Si attacca una nuova molecola di ATP.
Il ciclo viene quindi ripetuto fino a quando la concentrazione di ioni Ca 2+ diminuisce o la fornitura di ATP si esaurisce (a causa della morte cellulare). La velocità con cui la miosina scorre lungo l'actina è ≈15 μm/sec. Ci sono molte (circa 500) molecole di miosina nel filamento di miosina e, quindi, durante la contrazione, il ciclo viene ripetuto da centinaia di teste contemporaneamente, il che porta ad una contrazione rapida e forte. Va notato che la miosina si comporta come un enzima, un'ATPasi actina-dipendente. Poiché ogni ripetizione del ciclo è associata all'idrolisi dell'ATP, e quindi ad una variazione positiva dell'energia libera, il processo è unidirezionale. La miosina si muove lungo l'actina solo verso l'estremità positiva.
Fonte di energia per la riduzione
Per contrarre un muscolo viene utilizzata l'energia dell'idrolisi dell'ATP, ma la cellula muscolare dispone di un sistema estremamente efficiente per rigenerare l'apporto di ATP, per cui in un muscolo rilassato e in attività il contenuto di ATP è approssimativamente uguale. L'enzima fosfocreatinchinasi catalizza la reazione tra ADP e creatina fosfato, i cui prodotti sono ATP e creatina. La creatina fosfato contiene più energia immagazzinata dell'ATP. Grazie a questo meccanismo, durante l'esplosione di attività della cellula muscolare, il contenuto di creatina fosfato diminuisce, ma la quantità della fonte di energia universale - ATP - non cambia. I meccanismi di rigenerazione delle riserve di ATP possono variare a seconda della pressione parziale dell'ossigeno nei tessuti circostanti (vedi Organismi anaerobici).
Meccanismo normativo
Nella regolazione dell'attività muscolare sono coinvolti principalmente i neuroni, ma ci sono casi in cui gli ormoni (ad esempio l'adrenalina e l'ossitocina) controllano anche la contrazione della muscolatura liscia. Il segnale di contrazione può essere suddiviso in più fasi:
Dalla membrana cellulare al reticolo sarcoplasmatico
L'effetto di un trasmettitore rilasciato da un motoneurone provoca un potenziale d'azione sulla membrana cellulare della cellula muscolare, che viene ulteriormente trasmesso tramite speciali invaginazioni della membrana chiamate tubuli T, che si estendono dalla membrana nella cellula. Dai tubuli T, il segnale viene trasmesso al reticolo sarcoplasmatico, uno speciale compartimento di vescicole di membrana appiattite (reticolo endoplasmatico della cellula muscolare) che circonda ciascuna miofibrilla. Questo segnale provoca l'apertura dei canali del Ca 2+ nella membrana del reticolo. Gli ioni Ca 2+ posteriori entrano nel reticolo con l'aiuto di pompe di calcio a membrana - Ca 2+ -ATPasi.
Dal rilascio di ioni Ca 2+ alla contrazione delle miofibrille
Per controllare la contrazione, la proteina tropomiosina e un complesso di tre proteine - troponina (le subunità di questo complesso sono chiamate troponine T, I e C) sono attaccate al filamento di actina. La troponina C è uno stretto omologo di un'altra proteina, la calmodulina. C'è solo un complesso troponinico situato ogni sette subunità di actina. Il legame dell’actina alla troponina I sposta la tropomiosina in una posizione che interferisce con il legame della miosina all’actina. La troponina C si lega a quattro ioni Ca 2+ e indebolisce l'effetto della troponina I sull'actina e la tropomiosina occupa una posizione che non interferisce con la connessione dell'actina con la miosina. La fonte di energia per la contrazione delle fibre muscolari è l'ATP. Quando la troponina si lega agli ioni calcio, vengono attivati i centri catalitici per la scissione dell’ATP sulle teste della miosina. A causa dell'attività enzimatica delle teste della miosina, l'ATP situato sulla testa della miosina viene idrolizzato, il che fornisce energia per il cambiamento della conformazione delle teste e lo scorrimento dei filamenti. La molecola di ADP e il fosfato inorganico rilasciati durante l'idrolisi dell'ATP vengono utilizzati per la successiva risintesi dell'ATP. Una nuova molecola di ATP si attacca alla testa della miosina. In questo caso il ponte incrociato con il filamento di actina viene disconnesso. Il riattacco e il distacco dei ponti continuano finché la concentrazione di calcio all'interno delle miofibrille non diminuisce fino a raggiungere un valore sottosoglia. Quindi le fibre muscolari iniziano a rilassarsi.
Letteratura- B. Alberts, D. Bray, J. Lewis, M. Reff, K. Roberts, J. Watson, Biologia molecolare cellule - In 3 volumi - Trad. dall'inglese. - T.2. - M.: Mir, 1994. - 540 p.
- M. B. Berkinblit, S. M. Glagolev, V. A. Furalev, Biologia generale- In 2 ore - Parte 1. - M.: MIROS, 1999. - 224 p.: ill.
Che differiscono nell'organizzazione cellulare e tissutale, nell'innervazione e, in una certa misura, nei meccanismi di funzionamento. Allo stesso tempo, ci sono molte somiglianze nei meccanismi molecolari della contrazione muscolare tra questi tipi di muscoli.
Muscoli scheletrici
I muscoli scheletrici sono la parte attiva del sistema muscolo-scheletrico. Come risultato dell'attività contrattile dei muscoli striati, si verifica quanto segue:
- movimento del corpo nello spazio;
- movimento delle parti del corpo l'una rispetto all'altra;
- mantenendo una posa.
Inoltre, uno dei risultati della contrazione muscolare è la produzione di calore.
Nell'uomo, come in tutti i vertebrati, le fibre muscolari scheletriche hanno quattro importanti proprietà:
- eccitabilità- la capacità di rispondere ad uno stimolo mediante cambiamenti nella permeabilità ionica e nel potenziale di membrana;
- conduttività - la capacità di condurre un potenziale d'azione lungo l'intera fibra;
- contrattilità- la capacità di contrarsi o cambiare tensione quando eccitato;
- elasticità - la capacità di sviluppare tensione tensile.
In condizioni naturali, l'eccitazione e la contrazione muscolare sono causate dagli impulsi nervosi che entrano nelle fibre muscolari dai centri nervosi. Per provocare l'eccitazione in un esperimento, viene utilizzata la stimolazione elettrica.
La stimolazione diretta del muscolo stesso è detta stimolazione diretta; L'irritazione di un nervo motore che porta alla contrazione di un muscolo innervato da questo nervo (eccitazione delle unità neuromotorie) è un'irritazione indiretta. Dato che l'eccitabilità del tessuto muscolare è inferiore a quella del tessuto nervoso, l'applicazione di elettrodi di corrente irritante direttamente sul muscolo non fornisce ancora un'irritazione diretta: la corrente, diffondendosi attraverso il tessuto muscolare, agisce principalmente sulle terminazioni del motore nervi situati in esso e li eccita, il che porta alla contrazione dei muscoli.
Tipi di abbreviazione
Regime isotonico- una contrazione in cui il muscolo si accorcia senza creare tensione. Tale riduzione è possibile quando un tendine viene tagliato o rotto o in un esperimento su un muscolo isolato (rimosso dal corpo).
Modalità isometrica- una contrazione in cui la tensione muscolare aumenta, ma la lunghezza praticamente non diminuisce. Questa riduzione si osserva quando si tenta di sollevare un carico eccessivo.
Modalità auxotonica - una contrazione in cui la lunghezza di un muscolo cambia all'aumentare della sua tensione. Questa modalità di riduzione si osserva durante l’attività lavorativa di una persona. Se la tensione di un muscolo aumenta man mano che si accorcia, si chiama contrazione concentrico, e in caso di aumento della tensione muscolare durante l'allungamento (ad esempio, quando si abbassa lentamente un carico) - contrazione eccentrica.
Tipi di contrazioni muscolari
Esistono due tipi di contrazioni muscolari: singole e tetaniche.
Quando un muscolo viene irritato da un unico stimolo, si verifica un'unica contrazione muscolare, nella quale si distinguono le seguenti tre fasi:
- fase del periodo di latenza - inizia dall'inizio dello stimolo fino all'inizio dell'accorciamento;
- fase di contrazione (fase di accorciamento) - dall'inizio della contrazione al valore massimo;
- fase di rilassamento - dalla contrazione massima alla lunghezza iniziale.
Contrazione del singolo muscolo si osserva quando una breve serie di impulsi nervosi provenienti dai motoneuroni arriva al muscolo. Può essere indotto applicando uno stimolo elettrico molto breve (circa 1 ms) al muscolo. La contrazione muscolare inizia entro un intervallo di tempo fino a 10 ms dall'inizio dello stimolo, chiamato periodo di latenza (Fig. 1). Successivamente si sviluppano un accorciamento (durata circa 30-50 ms) e un rilassamento (50-60 ms). L'intero ciclo di una singola contrazione muscolare dura in media 0,1 s.
La durata di una singola contrazione nei diversi muscoli può variare notevolmente e dipende dallo stato funzionale del muscolo. La velocità di contrazione e soprattutto di rilassamento rallenta man mano che si sviluppa l'affaticamento muscolare. A muscoli veloci che hanno una singola contrazione a breve termine includono i muscoli esterni del bulbo oculare, delle palpebre, dell'orecchio medio, ecc.
Confrontando la dinamica della generazione di un potenziale d'azione sulla membrana della fibra muscolare e la sua singola contrazione, è chiaro che il potenziale d'azione si verifica sempre prima e solo allora inizia a svilupparsi un accorciamento, che continua dopo la fine della ripolarizzazione della membrana. Ricordiamo che la durata della fase di depolarizzazione del potenziale d'azione della fibra muscolare è di 3-5 ms. Durante questo periodo di tempo, la membrana fibrosa si trova in uno stato di assoluta refrattarietà, seguita dal ripristino della sua eccitabilità. Poiché la durata dell'accorciamento è di circa 50 ms, è ovvio che anche durante l'accorciamento la membrana della fibra muscolare dovrebbe ripristinare l'eccitabilità e sarà in grado di rispondere a un nuovo impatto con una contrazione sullo sfondo di uno incompleto. Di conseguenza, sullo sfondo dello sviluppo della contrazione nelle fibre muscolari, sulla loro membrana possono essere causati nuovi cicli di eccitazione e successive contrazioni cumulative. Questa riduzione cumulativa viene chiamata tetanico(tetano). Può essere osservato in una singola fibra e intero muscolo. Tuttavia, il meccanismo della contrazione tetanica in condizioni naturali in un muscolo intero ha le sue peculiarità.

Riso. 1. Relazioni temporali tra singoli cicli di eccitazione e contrazione delle fibre muscolari scheletriche: a - rapporto tra potenziale d'azione, rilascio di Ca 2+ nel sarcoplasma e contrazione: 1 - periodo di latenza; 2 - accorciamento; 3 - rilassamento; b - rapporto tra potenziale d'azione, eccitabilità e contrazione
Tetano chiamata contrazione muscolare che si verifica come risultato della somma delle contrazioni delle sue unità motorie causate dalla ricezione di numerosi impulsi nervosi dai motoneuroni che innervano questo muscolo. La somma delle forze sviluppate durante la contrazione delle fibre di più unità motorie aiuta ad aumentare la forza di contrazione del muscolo tetanico e influenza la durata della contrazione.
Distinguere frastagliato E liscio tetano. Per osservare il tetano dentato in un esperimento, il muscolo viene stimolato con impulsi di corrente elettrica ad una frequenza tale che ogni stimolo successivo viene applicato dopo la fase di accorciamento, ma prima della fine del rilassamento. La contrazione tetanica regolare si sviluppa con stimolazione più frequente quando vengono applicati stimoli successivi durante lo sviluppo dell'accorciamento muscolare. Ad esempio, se la fase di accorciamento del muscolo è di 50 ms, la fase di rilassamento è di 60 ms, quindi per ottenere il tetano seghettato è necessario irritare questo muscolo con una frequenza di 9-19 Hz, per ottenere il tetano liscio - con una frequenza di a almeno 20Hz.
Per dimostrazione vari tipi Il tetano di solito utilizza la registrazione grafica su un chimografo delle contrazioni del muscolo gastrocnemio isolato della rana. Un esempio di tale chimogramma è mostrato in Fig. 2.
Se confrontiamo le ampiezze e le forze sviluppate durante le diverse modalità di contrazione muscolare, sono minime con una singola contrazione, aumentano con il tetano seghettato e diventano massime con una contrazione tetanica regolare. Uno dei motivi di questo aumento dell'ampiezza e della forza di contrazione è che l'aumento della frequenza di generazione AP sulla membrana della fibra muscolare è accompagnato da un aumento della produzione e dell'accumulo di ioni Ca 2+ nel sarcoplasma delle fibre muscolari , che contribuisce ad una maggiore efficienza dell'interazione tra proteine contrattili.

Riso. 2. Dipendenza dell'ampiezza della contrazione dalla frequenza della stimolazione (la forza e la durata degli stimoli rimangono invariate)
Con un aumento graduale della frequenza di stimolazione, la forza e l'ampiezza della contrazione muscolare aumentano solo fino a un certo limite: la risposta ottimale. La frequenza di stimolazione che provoca la massima risposta muscolare è detta ottimale. Un ulteriore aumento della frequenza di stimolazione è accompagnato da una diminuzione dell'ampiezza e della forza di contrazione. Questo fenomeno è detto risposta pessima, mentre le frequenze di stimolazione che superano il valore ottimale sono dette pessimali. I fenomeni di ottimo e pessimo furono scoperti da N.E. Vvedensky.
In condizioni naturali, la frequenza e la modalità di invio degli impulsi nervosi dai motoneuroni al muscolo assicurano il coinvolgimento asincrono nel processo di contrazione di un numero maggiore o minore (a seconda del numero di motoneuroni attivi) di unità motorie del muscolo e del somma delle loro contrazioni. La contrazione di un muscolo integrale del corpo è di natura quasi teganica liscia.
Per caratterizzare l'attività funzionale dei muscoli, vengono valutati il loro tono e la loro contrazione. Il tono muscolare è uno stato di tensione continua prolungata causato dalla contrazione asincrona alternata delle sue unità motorie. In questo caso l'accorciamento visibile del muscolo può essere assente poiché non tutti i muscoli sono coinvolti nel processo di contrazione, ma solo quelli unità motorie, le cui proprietà il modo migliore adattati al mantenimento del tono muscolare e la forza della loro contrazione asincrona non è sufficiente ad accorciare il muscolo. Vengono chiamate contrazioni di tali unità durante la transizione dal rilassamento alla tensione o quando si cambia il grado di tensione Tonico. Vengono chiamate contrazioni a breve termine accompagnate da cambiamenti nella forza e nella lunghezza muscolare fisico.
Meccanismo di contrazione muscolare
Una fibra muscolare è una struttura multinucleata circondata da una membrana e contenente un apparato contrattile specializzato -miofibrille(Fig. 3). Inoltre, i componenti più importanti della fibra muscolare sono i mitocondri, i sistemi di tubuli longitudinali - il reticolo sarcoplasmatico e un sistema di tubuli trasversali - Sistema a T.

Riso. 3. La struttura della fibra muscolare
L'unità funzionale dell'apparato contrattile di una cellula muscolare è sarcomero, La miofibrilla è costituita da sarcomeri. I sarcomeri sono separati l'uno dall'altro da placche a Z (Fig. 4). I sarcomeri nella miofibrilla sono disposti in sequenza, quindi le contrazioni dei capcomeri causano la contrazione della miofibrilla e l'accorciamento complessivo della fibra muscolare.

Riso. 4. Schema della struttura del sarcomero
Lo studio della struttura delle fibre muscolari al microscopio ottico ha rivelato le loro striature trasversali, dovute alla speciale organizzazione delle proteine contrattili delle protofibrille - actina E miosina. I filamenti di actina sono rappresentati da un doppio filamento attorcigliato a doppia elica con un passo di circa 36,5 nm. Questi filamenti sono lunghi 1 µm e hanno un diametro di 6-8 nm, il cui numero raggiunge circa 2000, e sono attaccati ad un'estremità alla piastra Z. Le molecole proteiche filamentose si trovano nelle scanalature longitudinali dell'elica di actina tropomiosina. Con un passo di 40 nm, una molecola di un'altra proteina si attacca alla molecola di tropomiosina - troponina.
La troponina e la tropomiosina svolgono (vedi Fig. 3) un ruolo importante nei meccanismi di interazione tra actina e miosina. Al centro del sarcomero, tra i filamenti di actina, ci sono spessi filamenti di miosina lunghi circa 1,6 µm. Al microscopio polarizzatore, quest'area è visibile come una striscia di colore scuro (a causa della birifrangenza) - disco A anisotropo. Al centro è visibile una striscia più chiara H. A riposo non ci sono filamenti di actina. Su entrambi i lati UN- il disco è luce visibile isotropico strisce - I-dischi formato da filamenti di actina.
A riposo, i filamenti di actina e miosina si sovrappongono leggermente in modo che la lunghezza totale del sarcomero sia di circa 2,5 μm. Con la microscopia elettronica al centro H-strisce rilevate Linea M - struttura che contiene i filamenti di miosina.
La microscopia elettronica mostra che ai lati del filamento di miosina ci sono delle sporgenze chiamate ponti trasversali. Secondo i concetti moderni, il ponte trasversale è costituito da una testa e un collo. La testa acquisisce una pronunciata attività ATPasi legandosi all'actina. Il collo ha proprietà elastiche ed è un'articolazione incernierata, quindi la testa del ponte trasversale può ruotare attorno al proprio asse.
L'uso della tecnologia moderna ha permesso di stabilire che si applica la stimolazione elettrica a un'area Z-piastra porta alla riduzione del sarcomero, mentre la dimensione della zona del disco UN non cambia, ma la dimensione delle strisce H E IO diminuisce. Queste osservazioni hanno indicato che la lunghezza dei filamenti di miosina non cambia. Risultati simili sono stati ottenuti quando il muscolo veniva allungato: la lunghezza intrinseca dei filamenti di actina e miosina non cambiava. Come risultato degli esperimenti, si è scoperto che l'area di reciproca sovrapposizione dei filamenti di actina e miosina è cambiata. Questi fatti hanno permesso a X. e A. Huxley di proporre la teoria dello scorrimento del filo per spiegare il meccanismo della contrazione muscolare. Secondo questa teoria, durante la contrazione, la dimensione del sarcomero diminuisce a causa del movimento attivo dei filamenti sottili di actina rispetto ai filamenti spessi di miosina.

Riso. 5. A - diagramma dell'organizzazione del reticolo sarcoplasmatico, dei tubuli trasversali e delle miofibrille. B - diagramma della struttura anatomica dei tubuli trasversali e del reticolo sarcoplasmatico in una singola fibra muscolare scheletrica. B - il ruolo del reticolo sarcoplasmatico nel meccanismo di contrazione del muscolo scheletrico
Durante il processo di contrazione della fibra muscolare, si verificano le seguenti trasformazioni:
conversione elettrochimica:
- generazione del PD;
- distribuzione della PD attraverso il sistema T;
- stimolazione elettrica della zona di contatto del sistema T e del reticolo sarcoplasmatico, attivazione di enzimi, formazione di inositolo trifosfato, aumento della concentrazione intracellulare di ioni Ca 2+;
trasformazione chemiomeccanica:
- interazione degli ioni Ca 2+ con troponina, cambiamento nella configurazione della tropomiosina, rilascio di centri attivi sui filamenti di actina;
- interazione della testa della miosina con l'actina, rotazione della testa e sviluppo della trazione elastica;
- scorrimento dei filamenti di actina e miosina l'uno rispetto all'altro, riduzione delle dimensioni del sarcomero, sviluppo di tensione o accorciamento della fibra muscolare.
Il trasferimento dell'eccitazione dal motoneurone alla fibra muscolare avviene utilizzando il mediatore acetilcolina (ACh). L'interazione dell'ACh con il recettore colinergico della placca terminale porta all'attivazione dei canali sensibili all'ACh e alla comparsa di un potenziale della placca terminale, che può raggiungere i 60 mV. In questo caso la zona della placca terminale diventa una fonte di corrente irritante per la membrana della fibra muscolare e nelle zone della membrana cellulare adiacenti alla placca terminale si verifica una PD che si diffonde in entrambe le direzioni ad una velocità di circa 3-5 m/s ad una temperatura di 36 °C. Pertanto, la generazione del PD è la prima fase contrazione muscolare.
Seconda faseè la propagazione della PD nella fibra muscolare attraverso il sistema trasversale di tubuli, che funge da collegamento tra la membrana superficiale e l'apparato contrattile della fibra muscolare. Il sistema G è in stretto contatto con le cisterne terminali del reticolo sarcoplasmatico di due sarcomeri vicini. La stimolazione elettrica del sito di contatto porta all'attivazione degli enzimi situati nel sito di contatto e alla formazione di inositolo trifosfato. L'inositolo trifosfato attiva i canali del calcio delle membrane delle cisterne terminali, il che porta al rilascio di ioni Ca 2+ dalle cisterne e ad un aumento della concentrazione intracellulare di Ca 2+ "da 10 -7 a 10 -5. Il set dei processi che portano ad un aumento della concentrazione intracellulare di Ca 2+ è l'essenza terza fase contrazione muscolare. Pertanto, nelle prime fasi, il segnale elettrico dell'AP viene convertito in uno chimico - un aumento della concentrazione intracellulare di Ca 2+, ad es. conversione elettrochimica(Fig. 6).
Quando la concentrazione intracellulare degli ioni Ca 2+ aumenta, questi si legano alla troponina, che modifica la configurazione della tropomiosina. Quest'ultimo si mescolerà nel solco tra i filamenti di actina; in questo caso si aprono aree sui filamenti di actina con cui possono interagire i ponti incrociati della miosina. Questo spostamento della tropomiosina è dovuto ad un cambiamento nella formazione della molecola proteica della troponina in seguito al legame del Ca 2+. Di conseguenza, la partecipazione degli ioni Ca 2+ al meccanismo di interazione tra actina e miosina è mediata attraverso la troponina e la tropomiosina. Così, quarta fase l'accoppiamento elettromeccanico è l'interazione del calcio con la troponina e lo spostamento della tropomiosina.
SU quinta tappa L'accoppiamento elettromeccanico si verifica quando la testa del ponte trasversale della miosina si attacca al ponte di actina, al primo di numerosi centri stabili localizzati in sequenza. In questo caso, la testa della miosina ruota attorno al proprio asse, poiché presenta diversi centri attivi che interagiscono in sequenza con i centri corrispondenti sul filamento di actina. La rotazione della testa porta ad un aumento della trazione elastica del collo del ponte crociato e ad un aumento della tensione. In ogni momento specifico dello sviluppo della contrazione, una parte delle teste dei ponti trasversali è in connessione con il filamento di actina, l'altra è libera, cioè c'è una sequenza della loro interazione con il filamento di actina. Ciò garantisce un processo di riduzione regolare. Nella quarta e quinta fase avviene una trasformazione chemiomeccanica.

Riso. 6. Processi elettromeccanici nel muscolo
La reazione sequenziale di connessione e separazione delle teste dei ponti trasversali con il filamento di actina porta allo scorrimento dei filamenti sottili e spessi l'uno rispetto all'altro e ad una diminuzione delle dimensioni del sarcomero e della lunghezza totale del muscolo, che è sesta fase. La totalità dei processi descritti costituisce l'essenza della teoria dello scorrimento del filo (Fig. 7).
Inizialmente si credeva che gli ioni Ca 2+ fungessero da cofattore per l'attività ATPasi della miosina. Ulteriori ricerche hanno confutato questa ipotesi. Nel muscolo a riposo, actina e miosina non hanno praticamente alcuna attività ATPasi. L'attaccamento della testa della miosina all'actina fa sì che la testa acquisisca l'attività dell'ATPasi.

Riso. 7. Illustrazione della teoria dei fili scorrevoli:
A. a - muscolo a riposo: A. 6 - muscolo in contrazione: B. a. b - interazione sequenziale dei centri attivi della testa della miosina con i centri del filamento attivo
L'idrolisi dell'ATP nel centro ATPasi della testa della miosina è accompagnata da un cambiamento nella conformazione di quest'ultima e dal suo trasferimento in un nuovo stato ad alta energia. Il riattacco della testa della miosina a un nuovo centro sul filamento di actina porta nuovamente alla rotazione della testa, che è fornita dall'energia immagazzinata in essa. In ogni ciclo di connessione e separazione della testa della miosina con l'actina, una molecola di ATP viene scissa per ponte. La velocità di rotazione è determinata dalla velocità di degradazione dell'ATP. È chiaro che le fibre fasiche veloci consumano significativamente più ATP per unità di tempo e trattengono meno energia chimica durante l’esercizio tonico rispetto alle fibre lente. Pertanto, nel processo di trasformazione chemiomeccanica, l'ATP fornisce la separazione della testa di miosina e del filamento di actina e fornisce energia per l'ulteriore interazione della testa di miosina con un'altra parte del filamento di actina. Queste reazioni sono possibili a concentrazioni di calcio superiori a 10 -6 M.
I meccanismi descritti di accorciamento delle fibre muscolari suggeriscono che il rilassamento richiede prima una diminuzione della concentrazione di ioni Ca 2+. È stato sperimentalmente dimostrato che il reticolo sarcoplasmatico ha un meccanismo speciale: una pompa del calcio, che restituisce attivamente il calcio ai serbatoi. La pompa del calcio è attivata dal fosfato inorganico, che si forma durante l'idrolisi dell'ATP. e l'approvvigionamento energetico per la pompa del calcio è dovuto anche all'energia generata durante l'idrolisi dell'ATP. Pertanto, l'ATP è il secondo fattore più importante, assolutamente necessario per il processo di rilassamento. Per qualche tempo dopo la morte, i muscoli rimangono morbidi a causa della cessazione dell'influenza tonica dei motoneuroni. Quindi la concentrazione di ATP diminuisce al di sotto del livello critico e scompare la possibilità di separazione della testa della miosina dal filamento di actina. Il fenomeno del rigor mortis si verifica con pronunciata rigidità dei muscoli scheletrici.
Significato funzionale dell'ATP durante la contrazione del muscolo scheletrico- Idrolisi dell'ATP da parte della miosina, a seguito della quale i ponti trasversali ricevono energia per lo sviluppo della forza di trazione
- Legame dell'ATP alla miosina, che porta al distacco dei ponti trasversali attaccati all'actina, creando la possibilità di ripetere il ciclo della loro attività
- Idrolisi dell'ATP (sotto l'azione di Ca 2+ -ATPasi) per il trasporto attivo di ioni Ca 2+ nelle cisterne laterali del reticolo sarcoplasmatico, riducendo il livello di calcio citoplasmatico al livello iniziale

Somma delle contrazioni e del tetano
Se in un esperimento due forti stimolazioni singole agiscono su una singola fibra muscolare o su un intero muscolo in rapida successione, le contrazioni risultanti avranno un'ampiezza maggiore della contrazione massima durante una singola stimolazione. Gli effetti contrattili provocati dalla prima e dalla seconda irritazione sembrano sommarsi. Questo fenomeno è chiamato somma delle contrazioni (Fig. 8). Si osserva con irritazione muscolare sia diretta che indiretta.
Perché avvenga la sommatoria è necessario che l'intervallo tra le irritazioni abbia una certa durata: deve essere più lungo del periodo refrattario, altrimenti non si avrà risposta alla seconda irritazione, e più breve dell'intera durata della risposta contrattile, quindi che la seconda irritazione colpisce il muscolo prima che abbia il tempo di rilassarsi dopo la prima irritazione. In questo caso sono possibili due opzioni: se la seconda stimolazione arriva quando il muscolo ha già iniziato a rilassarsi, allora sulla curva miografica l'apice di questa contrazione sarà separato dall'apice della prima mediante retrazione (Figura 8, G-D) ; se la seconda stimolazione agisce quando la prima non ha ancora raggiunto il suo picco, allora la seconda contrazione si fonde completamente con la prima, formando un unico picco sommato (Figura 8, A-B).
Considera la sommatoria nel muscolo gastrocnemio della rana. La durata della fase ascendente della sua contrazione è di circa 0,05 s. Pertanto, per riprodurre su questo muscolo il primo tipo di sommatoria delle contrazioni (somma incompleta), è necessario che l'intervallo tra la prima e la seconda stimolazione sia superiore a 0,05 s, e ottenere il secondo tipo di sommatoria (la cosiddetta somma completa) - meno di 0,05 s.

Riso. 8. Somma delle contrazioni muscolari 8 risposta a due stimoli. Timestamp 20ms
Con la somma completa e incompleta delle contrazioni, i potenziali d'azione non vengono riassunti.
Muscolo del tetano
Se una singola fibra muscolare o l'intero muscolo viene sottoposto a stimolazione ritmica con una frequenza tale da sommare i loro effetti, si verifica una contrazione forte e prolungata del muscolo, detta contrazione tetanica, O tetano.
La sua ampiezza può essere molte volte maggiore della massima contrazione singola. Con una frequenza relativamente bassa di irritazione, si osserva tetano seghettato, A alta frequenza -tetano liscio(Fig. 9). Nel tetano le risposte contrattili del muscolo sono riassunte, ma le sue reazioni elettriche - potenziali d'azione - non sono riassunte (Fig. 10) e la loro frequenza corrisponde alla frequenza della stimolazione ritmica che ha causato il tetano.
Dopo la cessazione dell'irritazione tetanica, le fibre si rilassano completamente, la loro lunghezza originale viene ripristinata solo dopo un po 'di tempo. Questo fenomeno è chiamato contrattura post-tetanica o residua.
Quanto più velocemente le fibre muscolari si contraggono e si rilassano, tanto più frequente deve essere la stimolazione per provocare il tetano.
Affaticamento muscolare
L'affaticamento è una diminuzione temporanea delle prestazioni di una cellula, di un organo o di un intero organismo che si verifica a seguito del lavoro e scompare dopo il riposo.

Riso. 9. Tetano della fibra muscolare isolata (secondo F.N. Serkov):
a — tetano seghettato con una frequenza di stimolazione di 18 Hz; 6 - tetano liscio con una frequenza di stimolazione di 35 Hz; M - miogramma; P — segno di irritazione; B - marca temporale 1 s

Riso. 10. Registrazione simultanea della contrazione (a) e dell'attività elettrica (6) del muscolo scheletrico del gatto durante la stimolazione del nervo tetanico
Se irriti a lungo un muscolo isolato con stimoli elettrici ritmici, a cui viene sospeso un piccolo carico, l'ampiezza delle sue contrazioni diminuisce gradualmente fino a zero. Il record di contrazione registrato in questo caso è chiamato curva di fatica.
Una diminuzione delle prestazioni di un muscolo isolato durante un'irritazione prolungata è dovuta a due ragioni principali:
- Durante la contrazione, nel muscolo si accumulano prodotti metabolici (acido fosforico, lattico, ecc.) che hanno un effetto deprimente sulle prestazioni delle fibre muscolari. Alcuni di questi prodotti, così come gli ioni potassio, si diffondono dalle fibre nello spazio pericellulare e hanno un effetto deprimente sulla capacità della membrana eccitabile di generare potenziali d'azione. Se un muscolo isolato posto in un piccolo volume di liquido di Ringer viene irritato per lungo tempo e portato al punto di completo affaticamento, allora basta cambiare la soluzione lavandolo per ripristinare le contrazioni muscolari;
- progressivo deperimento nel muscolo riserve energetiche. Con il lavoro prolungato di un muscolo isolato, le riserve di glicogeno diminuiscono drasticamente, a seguito della quale il processo di risintesi di ATP e creatina fosfato, necessario per la contrazione, viene interrotto.
LORO. Sechenov (1903) ha dimostrato che il ripristino delle prestazioni dei muscoli stanchi del braccio di una persona dopo un lavoro prolungato di sollevamento di un carico viene accelerato se il lavoro viene svolto con l’altra mano durante il periodo di riposo. Il ripristino temporaneo della capacità lavorativa dei muscoli di un braccio stanco può essere ottenuto con altri tipi di attività motoria, ad esempio quando si lavorano i muscoli arti inferiori. In contrasto con il riposo semplice, tale riposo fu chiamato da I.M. Sechenov attivo. Considerava questi fatti come una prova che la fatica si sviluppa principalmente nei centri nervosi.
E hai già un'idea di cosa sia un muscolo. Ma come avviene la contrazione muscolare? Cosa fa funzionare i nostri muscoli?
parlando in un linguaggio semplice, la contrazione muscolare avviene sotto l'influenza degli impulsi nervosi che attivano le cellule nervose nel midollo spinale - neuroni motori, i cui rami sono assoni portato al muscolo. Se guardi più da vicino, all'interno del muscolo l'assone si divide e forma una rete di rami che, come contatti elettrici, sono “collegati” alla cellula muscolare. Attraverso tali contatti si verifica la contrazione muscolare.
Si scopre che ciascun motoneurone controlla un gruppo di cellule muscolari. Tali gruppi erano chiamati: unità neuromotorie, grazie al quale una persona può utilizzare parte del muscolo nel lavoro. Pertanto, possiamo controllare consapevolmente la velocità e la forza della contrazione muscolare.
Quindi, abbiamo esaminato il processo di “lancio” della contrazione muscolare. Ora diamo uno sguardo più da vicino a ciò che accade direttamente all'interno del muscolo durante la contrazione. Questo materiale è alquanto difficile da capire, ma molto importante. Devi capirlo, altrimenti non sarai in grado di comprendere appieno come crescono i nostri muscoli.
Contrazione muscolare in approssimativa approssimazione
Innanzitutto è necessario capire cosa è costituito da numerosi filamenti di due proteine: miosina E actina, che si trovano lungo la miofibrilla. Inoltre, la miosina è costituita da filamenti spessi e l'actina è costituita da filamenti sottili. Questo spiega la struttura a strisce chiaro-scure della miofibrilla (strisce scure - miosina, strisce chiare - actina).
In letteratura, le aree scure della miofibrilla sono chiamate disco A e le aree chiare sono chiamate disco I. I filamenti di actina sono attaccati alla cosiddetta linea Z, che si trova al centro del disco I. Viene chiamato il segmento della miofibrilla tra le linee Z, compreso il disco A della miosina sarcomero, che può essere considerata una sorta di unità contrattile della miofibrilla.
Il sarcomero si contrae come segue: con l'aiuto di rami laterali (ponti), i filamenti spessi di miosina trascinano su se stessi sottili filamenti di actina.

Cioè, le teste dei ponti si agganciano al filamento di actina e lo tirano tra i filamenti di miosina. Al termine del movimento le testine si disimpegnano e si agganciano nuovamente, continuando a ritrarsi. Si scopre che la contrazione muscolare è una combinazione di contrazioni di molti sarcomeri.
Se consideriamo separatamente il sottile filamento di actina, si tratta di una doppia elica di filamenti di actina, tra i quali si trova una doppia catena di tropomiosina.

Tropomiosinaè anche una proteina che blocca l'impegno dei ponti della miosina con l'actina in uno stato muscolare rilassato. Non appena un impulso nervoso viene fornito al muscolo attraverso un motoneurone, la polarità di carica della membrana della cellula muscolare cambia, per cui la cellula viene saturata con ioni di calcio (Ca++), che vengono rilasciati da depositi speciali situati lungo ciascuna miofibrilla. Il filamento di tropomiosina, in presenza di ioni calcio, si approfondisce istantaneamente tra i filamenti di actina e i ponti di miosina sono in grado di impegnarsi con l'actina: diventa possibile la contrazione muscolare.

Tuttavia, dopo che il Ca++ entra nella cellula, ritorna immediatamente al suo deposito e si verifica il rilassamento muscolare. Solo con impulsi costanti provenienti dal sistema nervoso possiamo mantenere una contrazione prolungata: questa condizione è stata definita contrazione muscolare tetanica.
Naturalmente, contrarre i muscoli richiede energia. Da dove viene, come si forma l'energia che sostiene il movimento del ponte di miosina? Lo imparerai nel prossimo articolo.
I materiali contenuti in questo articolo sono protetti dalla legge sul copyright. È VIETATO copiare senza fornire un collegamento alla fonte e avvisare l'autore!
- In contatto con 0
- Google+ 0
- OK 0
- Facebook 0






